Происхождение и эволюция органического мира
Страница 5
6) мутационная изменчивость не связана непосредственно с модификациями и независима от них;
7) мутации происходят почти во всех возможных направлениях;
8) способность к мутации наступает периодически.
И, наконец, пятое принципиальное положение, привнесенное генетикой, - реабилитация понятия вида как такового: вид – не удобная выдумка биологов, а реальная сущность, имеющая достаточно четкие границы и характеризующаяся собственным набором морфофизиологических признаков (фенотипом).
Из изложенного совершенно очевидно, что взгляды родоначальников генетики на движущие факторы эволюционного процесса, в особенности на изменчивость и наследственность, существенно отличны от воззрений Ламарка и Дарвина.
Тем не менее в 30-50-е годы трудами Добжанского, Симпсона, Майра и других была предпринята попытка примирить генетику с дарвинизмом. Так появилась синтетическая теория эволюции (СТЭ), создатели которой стремились обобщить все накопленные генетикой факты и в приложении к эволюционному учению объединить их с позиций Дарвина.
Основа построения синтетической теории эволюции – закон Харди – Вайнберга. Он наглядно показал, что благодаря синтезу с генетикой эволюционное учение постепенно утратило описательный характер и превратилось в математизированную теорию.
Знакомству с законом должно предшествовать краткое вступление о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира.
Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До конца 20-х годов текущего столетия данные генетики противопоставлялись эволюционной теории, наследственная изменчивость (мутационная, комбинативная) рассматривалась в качестве главного фактора эволюции, естественному отбору отводилась второстепенная роль. Таким образом, уже в начальный период своего становления генетика была использована для создания новых концепций эволюции. Сам по себе этот факт знаменателен: он свидетельствовал о тесной связи генетики с эволюционной теорией, но время их объединения было еще впереди. Различного рода критика дарвинизма была широко распространена вплоть до возникновения СТЭ.
Исключительную роль в развитии эволюционного учения сыграла популяционная генетика, исследующая микроэволюционные процессы в природных популяциях. Основана она выдающимися отечественными учеными С.С. Четвериковым и Н.В. Тимофеевым-Ресовским.
Начавшееся в 20-х годах объединение дарвинизма и генетики способствовало расширению и углублению синтеза дарвинизма с другими науками. 30 – 40-е годы принято считать периодом становления синтетической теории эволюции.
В западных странах обновленный дарвинизм, или синтетическая теория эволюции, приобрел широкое признание среди ученых уже в 40-х годах, хотя всегда были и есть отдельные крупные исследователи, занимающие антидарвиновские позиции.
Основные положения СТЭ выводятся как следствия из закона Харди-Вайнберга. Известно, что понимание сущности и значения закона вызывает у школьников затруднение, хотя его математический аппарат прост и доступен всем, кто знаком с алгеброй средней школы. Важно сосредоточить внимание учащихся не только на определении закона – частоты генов и генотипов в популяции не меняются в ряду поколений, - его условиях – бесконечно большая популяция, случайное свободное скрещивание особей, отсутствие мутационного процесса, естественного отбора и других факторов, - математической модели – AA p2 + Aa 2 p + aaq2 = 1, - но и на практическом применении закона.
Процесс микроэволюции доступен непосредственному изучению, о его наличии можно судить по изменяющемуся генофонду. Генофонд описывается либо частотами аллелей, либо частотами генотипов, соотношение которых легко установить. Для этого нужно подсчитать гомозиготных носителей рецессивного признака и, пользуясь простой формулой, вычислить частоту встречаемости аллелей, доминантных гомозигот и гетерозигот. Зная соотношение генотипов и аллелей в данной популяции, можно проследить ее дальнейшую эволюционную судьбу. Например, один из 10000 человек альбинос, т.е. частота гомозигот по рецессивному аллелю (q2) составляет 0,0001. Зная, что q2 = 0, 0001, можно определить частоту аллеля альбинизма (q), доминантного аллеля нормальной пигментации (p), гомозиготного доминантного генотипа (p2), гетерозиготного генотипа (2 pq):
p2 = 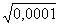 = 0,01 (или 1%).
= 0,01 (или 1%).
Поскольку p = 1 – q, то 1 – 0, 01 = 0,99; частота доминантного аллеля в популяции равна 0,99, или 99%. Нормально пигментированным людям будут соответствовать два генотипа (АА, Аа). Гомозиготный доминантный генотип будет встречаться с частотой 0,98, так как p2 = 0,992 = 0,98. Частота встречаемости гетерозигот составляет примерно 0,02, или 2%, так как 2pq = 2 x 0, 99 х х 0,01 = 0,0198 = 0,02.
Важно обратить внимание на следствия, вытекающие из закона:
рецессивные аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном. Состоянии. (Как видно из рассмотренного примера, при малом числе альбиносов, 200 человек из 10000 (2%) несут аллель альбинизма в скрытом, гетерозиготном состоянии);
в отличие от идеальной популяции на популяцию в природе оказывают давление эволюционные факторы, которые приводят к изменению ее генофонда.
В процессе дальнейшего изложения материала следствия закона Харди-Вайнберга будут углубляться, конкретизироваться. Так, при рассмотрении роли изменчивости в эволюционном процессе, что именно исходя из закона Харди-Вайнберга отечественный генетик С.С. Четвериков первым высказал мысль о значении рецессивных мутаций: они не исчезают в популяции, не растворяются в массе нормальных особей, а поглощаются свободным скрещиванием, оставаясь скрытыми в гетерозиготном состоянии в недрах вида, и проявляются фенотипически только в популяции. С работы С.С Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) и начался период синтеза генетики и эволюционного учения. Ученый сделал важный вывод о насыщенности природных популяций большим количеством рецессивных мутаций. Существование такого скрытого резерва наследственной изменчивости создает возможность для эволюционных преобразований в популяции под воздействием естественного отбора.
Таким образом, мы подводим к пониманию положений синтетической теории эволюции о том, что микроэволюционные процессы начинаются в популяции; материалом для эволюции служат чаще всего рецессивные мутации.
Другое следствие закона раскрывается при изучении эволюционных факторов. Условия идеальной популяции не соблюдаются в природе. Идеальная популяция не тождественна реальной. Под давлением эволюционных факторов частоты аллелей и генотипов в популяции изменяются, и это является основой всех эволюционных преобразований. Понятие каждого конкретного фактора эволюции вводится как результат поочередной замены условий идеальной популяции на противоположные. Например, если постоянство частот аллелей и генотипов в идеальной популяции имеет силу при бесконечно большой численности особей, то в малой природной популяции случайные процессы могут привести к заметным последствиям, в частности, к изменению ее генофонда. Такое противопоставление позволяет логично перейти к рассмотрению дрейфа генов – ненаправленного случайного изменения частот аллелей в малых популяциях, одна из причин которого – популяционные волны.
И еще. В идеальной популяции все генотипы одинаково плодовиты, т.е. естественного отбора не происходит, на реальную же популяцию естественный отбор оказывает существенное давление. В связи с этим раскрывается сущность естественного отбора как направляющего эволюционного фактора, выявляются его формы, творческая роль.