Пространственная ориентация живых организмов посредством зрительной сенсорной системы
Страница 5
1) спектр поглощения отдельного рабдомера хорошо совпадает: а) с известной кривой Дартнелла (Dartnall, 1953), б) со спектром поглощения родопсина быка и кальмара, в) с кривой спектральной чувствительности, измеренной по электроретинограмме, г) с кривой спектральной чувствительности, полученной при внутриклеточном отведении потенциалов;
2) спектры поглощения рабдомеров неодинаковы: так, шесть рабдомеров одной и той же ретинулы имеют максимум поглощения при 500 нм, а один — центральный — при 460 нм, что, вероятно, соответствует «синечувствительным» и «зеленочувствительным» рецепторам Буркхарда (Burkhardt, 1962);
3) после длительного засвета величина максимума поглощения при 500 нм уменьшается; вероятно, это является результатом частичного обесцвечивания зрительного пигмента, подобно тому как это имеет место у позвоночных;
4) рабдомерам свойственно дихроичное поглощение света, причем коэффициент дихроичности поглощения равен для Calliphora 4/3, т. е. 1.33.
Последний факт представляет, пожалуй, наибольший интерес, поскольку он позволяет вплотную приблизиться к вопросу об ориентации молекул зрительного пигмента в микровиллах рабдомеров и заслуживает более подробного рассмотрения.
Прежде всего следует вспомнить, что все животные, обладающие рабдомерным типом сетчатки (т. е. членистоногие и головоногие), способны ориентироваться в пространстве по плоскости поляризации света. В 1950 г. Аутрумом и Штумпфом была выдвинута гипотеза о том, что одиночная зрительная клетка может служить анализатором поляризованного света (Autrum
a. Stumpf, 1950), а вскоре благодаря развитию электронной микроскопии были расшифрованы особенности структуры, осуществляющей эту функцию, т.е. рабдомера (Fernandez-Moran, 1956; Goldsmith a. Philpott, 1957). Появились многочисленные работы, подтверждающие реакцию одиночной зрительной клетки членистоногих на поворот плоскости поляризации падающего света [мухи Lucilia (Burkhardt u. Wendler, 1960), мухи Calliphora (Autrum u. Zwebl, 1962a), краба Carcinus maenas и саранчи Locusta (Shaw, 1966, 1967) 1. Стало ясно, что если ретинулярная клетка способна анализировать плоскополяризованный свет, то непосредственным анализатором его должен являться рабдомер. В свою очередь рабдомер может служить анализатором плоскополяризованного света только при наличии дихроичности коэффициента поглощения (Vries, Spoor, Jielof, 1953; Stockbammer, 1956). В 1961 г.Муди и Паррисс (Moody a. Parriss, 1961), обнаружившие, что осьминог способен различать плоскость поляризации света, попытались оценить величину коэффициента дихроичности рабдомера. При этом они исходили из предположения, что зрительный пигмент является структурным элементом мембраны микровилл рабдомеров, подобно тому как это имеет место у позвоночных (Brown, Gibbons a. Wald, 1963). Более того, Мудн и Паррисс считали (тоже по аналогии с позвоночными), что дипольные моменты молекул зрительного пигмента лежат в плоскости мембраны микровилл, но ориентированы в ней хаотично (Wald, Brown a. Gibbons, 1963). В результате довольно несложного анализа Муди и Паррисс получили для относительного поглощения одиночной микровиллы выражение
|
| 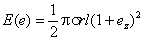 |
где Е – относительное поглощение света; s — поверхностная плотность молекул зрительного пигмента в мембране микровиллы (число молекул на единицу площади поверхности мембраны мпкровиллы); r – радиус микровиллы; l — длина микровиллы;
е — единичный вектор напряженности электрического поля, имеющий составляющие по трем осям координат, соответственно еx, еy и ez (в расчетах Муди и Паррисса считалось, что ось микровиллы совпадает по направлению с осью z).
Если свет распространяется вдоль оси у, которая параллельна оптической оси омматидия, то он имеет две составляющие электрического вектора — еx и ez . Дихроизм поглощения определяется отношением относительных поглощений для двух составляющих еx и ez , а в случае опытов с поворотом плоскости поляризации света для двух положений вектора е, т. е. когда он параллелен оси z, (или, что то же самое, оси микровиллы) – Е|| и когда он перпендикулярен ей – Е^.
Очевидно, что для е, параллельного ocи z, получается |e| = ez = 1, и равенство (1) дает
Соответственно для е, перпендикулярного оси z (т. е. для е, параллельного оси х), имеем
|e| = ex = 1, а ez = 0;
тогда
Коэффициент дихроичности поглощения (который по определению больше единицы) определяется отношением большего относительного поглощения (Е||) к меньшему (Е^), т. е. равен:
что и было получено Муди и Парриссом.
Однако значение коэффициента дихроичностп поглощения, полученное Лангером и Тореллом (Langer a. Thorell, 1966), меньше 2, а именно 4/3. Возможно, что причина этого заключается в том, что не все молекулы зрительного пигмента лежат в плоскости мембраны, что, как показали Уолд, Браун и Джиббонс, имеет место для мембран дисков наружных сегментов позвоночных (Wald, Brown a. Gibbons, 1963). В этом случае в расчеты Муди и Паррисса Грибакин (1969) предлагает внести поправку. Согласно этой поправке, следует считать, что дипольные моменты части молекул зрительного пигмента ориентированы перпендикулярно поверхности мембраны. Обозначим поверхностную плотность молекул с дипольными моментами, лежащими в плоскости мембраны, через ss, а поверхностную плотность молекул с дипольными моментами, ориентированными перпендикулярно поверхности мембраны, через sr (будем называть их радиальными дипольными моментами, так как эти моменты направлены по радиусам микровилл). Очевидно, что общая поверхностная плотность молекул зрительного пигмента равна s = ss + sr . При этом различным положениям плоскости поляризации световой волны (т. е. различным ориентациям вектора е, который перпендикулярен плоскости поляризации и лежит в так называемой плоскости колебаний электромагнитной волны) соответствуют и различные относительные поглощения молекул, входящих в ss , и молекул, входящих в sr. Так, для е, параллельного оси z, получим
Es^ = pssrl, Er|| = 0
(Еr || равна нулю, так как е параллельно оси z и, следовательно, перпендикулярно плоскости ХОУ, параллельно которой лежат радиально направленные моменты).
Для
е, перпендикулярного оси z, получим