Старение
Страница 4
Коллаген, выделенный из различных органов старых животных, резко отличается от коллагена, выделенного из тех же органов молодых. Эти изменения разнообразны, однако большинство из них является следствием одних и тех же молекулярных событий — образования между цепями коллагена ковалентных сшивок (чаще всего между их аминокислотами: лизином и тирозином). Количество таких сшивок столь закономерно увеличивается по мере старения, что физико-химические свойства коллагена могут служить критерием биологического возраста организма. Именно биологического, а не календарного. Например, после хронического облучения животных ионизирующей радиацией продолжительность жизни их уменьшается, причем в органах таких животных развиваются изменения, аналогичные тем, которые можно наблюдать и при старении тех же животных. То есть облучение ускоряет развитие процесса старения на уровне целостного организма. Развитие спонтанных молекулярных изменений межклеточного вещества также ускоряется: коллаген, выделенный из органов облученных животных, содержит количество сшивок, характерное для коллагена более старых животных.
В межклеточном веществе, кроме сети коллагеновых волокон, содержатся эластические нити. Они состоят из эластина, образование которого происходит в результате взаимодействия двух макромолекул: гликопротеида и проэластина. Причем сначала гликопротеид полимери-зуется в микрофибриллы, на которые в результате электростатического взаимодействия адсорбируются молекулы проэластина. Следующий этап образование эластиновых фибрилл катализируется специальным ферментом лизиноксидазой. В результате между двумя макромолекулами образуются ковалентные cвязи. Эластин также является метаболически стабильной структурой, и с возрастом количество перекрестных связей в нем увеличивается. Однако, образование этих «дополнительных» сшивок| происходит, очевидно, не из-за специфического действия ферментов, а вследствие окисления лизиновых остатков перекисями и другими продуктами метаболизма.
Таким образом, основное принципы старения клеток и межклеточного вещества оказываются одинаковыми. В обоих случаях они обусловлены образованием сшивок между макромолекулами, как правило, не подверженных метаболизму. Причем механизмы сшивания макромолекул также аналогичны. Процесс обычно начинается (инициируется) истинно спонтанным, тепловым разрушением макромолекул или окислением ее низкомолекулярными метаболитами (обычно перекисями), и при образовании сшивок между ДНК и гистонами и между молекулами коллагена и макромолекулами, входящими в состав эластина, во всех случаях, очевидно, участвуют аминогруппы лизина.
Мы уже отмечали, что молекулярные изменения межклеточного вещества имеют существенное значение в нарушении при старении взаимодействия между клетками, нарушении их снабжения кислородом и различными питательными веществами. Это связано не только, с тем, что затрудняется диффузия веществ между клетками, а также между ними и кровью. Ведь и коллагеновые, и эластиновые волокна входят в состав сосудистой стенки. Следовательно, то, что последняя с возрастом становится более плотной, менее эластичной, зависит прежде всего от рассмотренных нами молекулярных изменений коллагена и эластина. Более того, такие изменения являются тем основным фактором, .который делает сосудистую стенку с возрастом более уязвимой к атеросклерозу. Это конкретный пример того, что старение подготавливает почву для развития наиболее распространенных и тяжелых заболеваний человека.
Однако старение межклеточного вещества, хотя и может инициироваться независимо от возрастных изменений клеток, тем не менее в значительной степени определяется ими. Митотический потенциал фибробластов с возрастом уменьшается. Но при старении нарушается не только этаих функция, но и способность синтезировать коллаген.
[7]Образование эластиновых волокон зависит от координированного синтеза клетками по крайней мере двух типов макромолекул. В «старой» соединительной ткани эта координация нарушается, синтезируется относительно большее количество- гликопротеида, что, естественно, должно приводить образованию эластиновых нитей с нарушенной структурой, а это, в свою очередь, служит основой для возникновения «противоэластиновых» антител, разрушающих стенку сосуда. Такие изменения происходят не только в процессе «нормального» старения сосудистой стенки, но и составляют цепь событий при развитии атеросклероза. Даже старение межклеточного вещества костной ткани, вещества исключительно инертного, очевидно, в значительной степени зависит от нарушения функции специальных клеток — остеобластов. Именно эти клетки синтезируют вещество (белок, находящийся в комплексе с кальцием и фосфатом), которое инициирует процесс отложения в кости солей кальция. При различных патологических процессах отложение кальция происходит не только в костной ткани, но и других органах (чаще всего в артериальной стенке). Причиной этого процесса — дистрофической кальцификации является не увеличение в крови концентрации кальция, а скорее нарушение синтеза остеобластами инициатора кальцификации
Мы кратко остановились на рассмотрении механизма этого явления, потому что дистрофическая кальцификация (окостение различных тканей) — один из важных симптомов старения человека. Следовательно, и этот, казалось бы, чисто физико-химический процесс изменения с возрастом межклеточного вещества может быть связан со старением клеток.
5. Проявление старения на тканевом уровне.
[8]О существовании внутриклеточных факторов, ограничивающих продолжительность жизни клеток пределами, характерными для данного вида, свидетельствую и с исследования проведённые на культуре ткани.
Основатель метода Carrel в 1912г. ввёл в культуру ткани фибробласты сердечной мышцы зародыша цыплёнка путём непрерывных пассажей в свежую питательную среду фибробласты сохранялись в течении 34л. (Parken, 1961г.). Клетки всё время были молодыми и здоровыми и прожили тройной срок жизни цыплёнка, эквивалентной 200 годам жизни человека. Опыт был закончен в 1946г.
[9]С целью исследования причин старения организма было предпринято изучение старения клеток в культуре. Как впервые показали Свим и Паркер, фибробласты, полученные из различных тканей человека, пролиферируют только в течение ограниченного времени, после чего погибают. Эти авторы, однако, не выяснили, сохраняют ликлетки в культуре свои нормальные свойства, и не осмелились предположить, что их ограниченная пролиферативная активность связана со старением. Позже аналогичные работы были широко развернуты в лаборатории Хейфлика. Хейфлик и Мурхед установили, что легочные фибробласты эмбриона человека проходят в культуре около 50 удвоений популяции и затем погибают. Фибробласты, полученные из легочной ткани взрослого донора, подвергаются только ~20 удвоениям популяции. Хейфлик предположил, что имеется обратная зависимость между возрастом донора и пролиферативным потенциалом фибробластов. Позже другие исследователи также сообщили о наличии такой связи 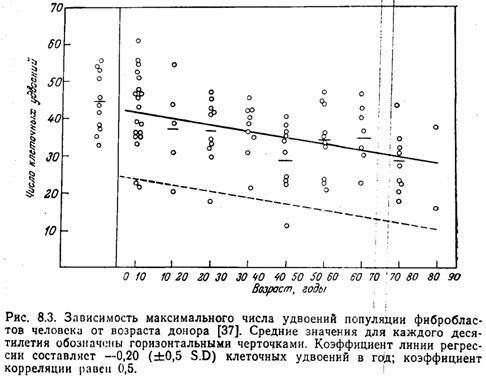
Более того, было показано, что при культивировании фибробластов кожи, полученных от людей в возрасте 10—90 лет, наблюдается снижение потенциала удвоения примерно на 0,20 относительные единицы в расчете на каждый год жизни донора. Коэффициент регрессии составляет —0,20. Сообщение Голдстейна и др. о том, что не хронологический возраст донора, а скорее его физиологическое состояние ответственно за скорость роста фибробластов, побудило многих авторов проделать скрупулезную работу с клетками разных типов, чтобы установить, действительно ли существует линейная связь между хронологическим возрастом и удвоением популяции. Изучение культур фибробластов, источником которых служили клетки тщательно отобранных здоровых доноров, не предрасположенных к диабету, дало отрицательный результат: обратная зависимость между возрастом донора и числом удвоений популяции не была обнаружена. В то же время эта зависимость была установлена для культур клеток, полученных от больных диабетом или от лиц, генетически предрасположенных к этому заболеванию. Таким образом, именно физиологическое состояние в большей степени, чем хронологический возраст, определяет пролиферативную активность клеток.