Математическое моделирование биосинтеза продуктов метаболизма
Страница 2
Рис. 2. Форма зависимости qР(m) для уравнений (13) и (14).
По аналогии с уравнениями (11) и (12) уравнение (14) начинается, не из 0 по оси ординат, а из некоторой точки a, что означает начало синтеза продукта без роста биомассы.
Субстрат-зависимые модели кинетики биосинтеза продуктов метаболизма.
С точки зрения математики, уравнения, в которых в качестве аргумента выступает m, предполагают, что совсем неважно, каким образом формируется то или иное значение m. Например, величину mможно изменятьпутем уменьшения концентрации углеродного субстрата или азотного субстрата, или путем снижения температуры или повышения величины рН. Для процессов, связанных только с ростом, возможно скорость биосинтеза при этом будет одинакова. Для несвязанных с ростом процессов небезразлично, каким путем мы будем изменять величину m. Лимитирование углеродом, лимитирование азотом, повышение рН или снижение температуры, давая одно и то же значение скорости роста, могут давать совершенно различные скорости биосинтеза продукта метаболизма. Другими словами, связь между qP и m не имеет строго причинно-следственного характера, а обусловлена влиянием на обе эти кинетические характеристики одних и тех же факторов внешней среды. Для таких процессов необходимо использовать уравнения, которые в качестве аргументов содержат независимо влияющие первичные факторы: концентрация того или иного субстрата, температура или величина рН.
Биосинтез продукта может описываться однофакторными или многофакторными уравнениями. Кроме того, было установлено, что структуры зависимостей qP от S, P, температуры и величины рН аналогичны структурам таких же уравнений для роста биомассы, например: Моно, Андрюса, Перта, Хиншельвуда и т.д. Например, если субстрат влияет на qP по Андрюсу, то имеем [10]:
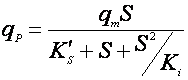 (15),
(15),
где qm – максимальная удельная скорость биосинтеза продукта;
K’S – константа насыщения;
Ki – константа ингибирования продуктом.
Многофакторные зависимости здесь чаще бывают мультипликативными, чем аддитивными. Приведем зависимость мультипликативного [11] и аддитивного влияния концентрации субстрата по механизму Моно:
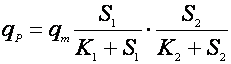 (16),
(16),
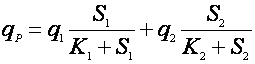 (17).
(17).
Применяются также уравнения с не разделяющимися эффектами факторов, например, типа Контуа [10] или неконкурентного торможения продуктом [5]:
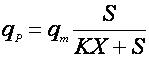 (18),
(18),
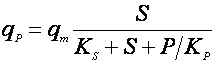 (19).
(19).
К сожалению, невозможно изложить все кинетические зависимости биосинтеза продуктов от первичных факторов в столь короткой работе. Данный материал подробно изложен в монографии [14], в которой приведены не только несколько десятков уравнений, но и произведен их анализ.
Модели, основанные на концепции возраста культуры микроорганизмов.
Для биосинтеза продуктов метаболизма часто бывает недостаточно только благоприятных “внешних” факторов среды. Потому что в биосинтезе участвуют внутриклеточные ферменты микроорганизмов, промежуточные продукты, содержание которых в клетке зависит от предыстории развития культуры. Слишком быстро выросшая культура часто неэффективна с точки зрения биосинтеза продукта. У микробиологов есть выражение “культура ушла в ботву”, что означает биомассы – много, продукта – мало или вообще нет. Однако, учитывать эти внутриклеточные компоненты при моделировании очень проблемно – их трудно измерять и соответственно находить кинетические коэффициенты.
Вместо этого предложены некоторые феноменологические подходы к оценке физиологического состояния микробной биомассы, основанные на оценке возрастного состояния популяции клеток.
Есть несколько подходов для учета возраста культуры. Один из них заключается в определении распределения клеток микроорганизмов по возрастам [12]. Тогда значение удельной скорости биосинтеза продукта можно считать как бы суммой скоростей, даваемых разными возрастными фракциями биомассы:
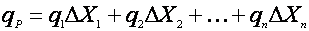 (20),
(20),
где ΔXi – концентрация биомассы i-ой возрастной группы;
qi – удельная скорость биосинтеза биомассой i-ой возрастной группы.
При этом, вполне возможно, что значения q1, q2, . ,qn не будут одинаковыми: “молодежь” не синтезирует нужный продукт, слишком старые клетки – тоже.
Японским ученым Аибой был предложен более простой подход, использовать для оценки возраста культуры так называемый средний возраст популяции 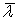 как параметр, определяющий биосинтетическую активность культуры [13]. Биологически термин вполне понятен – это сумма возрастов всех клеток, деленная на их количество:
как параметр, определяющий биосинтетическую активность культуры [13]. Биологически термин вполне понятен – это сумма возрастов всех клеток, деленная на их количество:
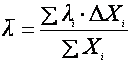 (21),
(21),
где λi – возраст i-ой возрастной группы.
Если последовательно уменьшать поддиапазоны Δt и ΔΧ, доведя их до бесконечно малых dX и dt, то для среднего возраста можно получить интегральную формулу:
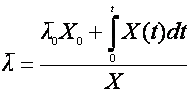 (22),
(22),
где Х0 – начальная концентрация биомассы;
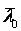 – средний возраст культуры в начальный момент культивирования.
– средний возраст культуры в начальный момент культивирования.
Другим способом упрощения возрастной зависимости является разделение возрастного диапазона клеток на 2 класса – продуктивный (выше некоторого значения) и не продуктивный [14]:
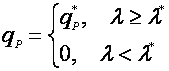 (24),
(24),
где λ* – возраст зрелости;
qP* – удельная скорость биосинтеза клетки, по достижении ею возраста зрелости.
Теперь остается рассмотреть форму зависимости удельной скорости биосинтеза продукта qР от среднего возраста культуры: 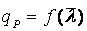 .
.
Если зависимость имеет возрастающий характер с насыщением, то зависимость удобно выразить в форме, похожей на уравнение Моно:
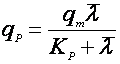 (25).
(25).
Если, наоборот, она падает с возрастом, то лучше подходит выражение, подобное уравнению Иерусалимского:
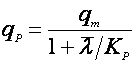 (26).
(26).
Если зависимость имеет экстремум, то оно может быть выражена, например, с помощью аппроксимирующего полиномиального уравнения [15]:
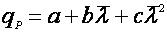 (27).
(27).
Однозначная зависимость между qР и на практике встречается редко, часто зависимость скорости биосинтеза продукта от возраста учитывают в виде мультипликативного сомножителя, сопряженного с основной частью уравнения, учитывающего влияния остальных факторов.
Модели деградации (инактивации) продуктов метаболизма.
Не всегда синтезированные продукты метаболизма остаются устойчивыми; часто они настолько нестабильны, что разрушаются уже в процессе самой ферментации. Поэтому, описывая материальный баланс по продукту метаболизма, необходимо учитывать кинетику его инактивации:
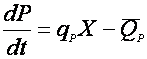 (28),
(28),
где 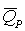 – скорость деградации продукта метаболизма.
– скорость деградации продукта метаболизма.
При рассмотрении синтеза метаболитов, использовалась удельная скорость, в случае деградации, вводить удельную скорость не корректно, т.к. продукт существует отдельно от биомассы, и его деградация не зависит в общем случае от ее концентрации.
Рассмотрим модели кинетики деградации:
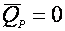 (29),
(29),
деградация отсутствует.
 (30),
(30),
деградация идет с постоянной скоростью. Такое выражение странно выглядит в начале процесса, когда продукта еще нет; из уравнения же получается, что концентрация продукта снижается ниже нуля, что не имеет физического смысла.