Метаболические сдвиги в организме, происходящие вследствие сахарного диабета
Страница 8
«сдерживающий» эффект инсулина на кетонемию обусловливается его способностью тормозить липолиз, снижать окисление жирных кислот до
кетоновых тел в печени и стимулировать утилизацию последних мышца-
ми. При тяжелой инсулиновой недостаточности увеличивается как до-ставка жирных кислот в печень, так и активность фермента, ограничива-
ющего скорость окисления жирных кислот в данном органе (ацилкарни-
тинтрансфераза). Изменения активности этого фермента в печени опосре-
дуется повышением содержания карнитина и снижением уровня малония -
КОА (первый, промежуточный продукт синтеза жирных кислот), который
в норме ингибирует ацилкарнитинтрансферазу.
1.2.5. НАРУШЕНИЕ БЕЛКОВОГО ОБМЕНА В РЕЗУЛЬТАТЕ
ПАТОЛОГИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ГОРМОНА.
Выраженный дефицит инсулина сопровождается отрицательным азотис-
тым балансом и резким белковым истощением. При ювенильном инсулин-
зависимом диабете частым осложнением в случае некомпенсированного заболевания является задержка роста. Такие нарушения не вызывают удивления, ибо инсулин, если он присутствует в нормальных количествах,
стимулирует синтез белка и поглощение аминокислот мышцами и тормо-зит расход белка и высвобождение аминокислот мышечной тканью. Изме-
нения белкового обмена сказываются и на глюконеогенезе, поскольку из-быточная продукция глюкозы при диабете, сопровождающемся кетозом отчасти зависит от повышения утилизации образующихся из белка пред-шественников.
При инсулинозависимом диабете с легко или умеренно выраженной ги-пергликемией изменяется содержание аминокислот в крови, их поглоще-ние печенью и высвобождение мышцами. При спонтанном диабете у чело-
века неоднократно отмечали снижение концентрации (аланина) в плазме
и повышение концентрации аминокислот. Несмотря на снижение уровня
аланина в плазме, поглощение этой глюкогенной аминокислоты и других
предшественников глюкозы печенью увеличивается в 2 раза и более
(Wahren J., 1972). Вследствие такого повышения поглощения субстратов на долю глюконеогенеза приходится более 30-40% от общей продукции
глюкозы печенью, тогда как у здорового человека эта величина составля-ет 15-20%. Поскольку содержание аланина в крови при диабете снижает-ся, увеличение его поглощения печенью обусловливается повышением
фракционной экстракции этой аминокислоты. В отсутствии нормального
«сдерживающего» эффекта инсулина на глюконеогенез печень выступает
в роли сифона, снижающего концентрацию аланина в артериальной кро-ви.
У больных диабетом количество азотистых продуктов в мышце пос-ле приема белковой пищи восстанавливается труднее, чем в норме. В от-
личие от интенсивного и длительного поглощения аминокислот с раветв-
ленной цепью мышичной тканью сопровождающее прием белковой пищи
у здорового человека, у больных диабетом наблюдается лишь транзитор-ное поглощение их. Вследствие этого снижается общее поглощение амино-
кислот мышцами, а уровень аминокислот с разветвленной цепью в плазме
после приема белковой пищи чрезмерно повышается (Wahren J. et all, 1976). Это согласуется с известным стимулирующим влиянием инсулина на поглощение мышцами аминокислот, особенно с разветвленной цепью
увеличение концентрации в артериальной крови, а снижение поглощения аминокислот после приема белковой пищи указывают на то, что диабет
характеризуется нарушением не только к глюкозе, но и к белку. Наруше-ния белкового обмена при диабете усугубляются тем, что аминокислоты,
захваченные мышечной тканью, не включаются в белок, а преимущест-венно распадаются (Felig P., 1985).
Торможение синтеза белка из аминокислот является предпосылкой для
образования из них углеводов. При сахарном диабете образование углево-
дов из белка, значительно увеличивается. Неоглюкогенез из белка возрас-
тает под влиянием АКТГ и глюкокартикоидов.
Изменение нейроэндокринной регуляции обменных процессов приводит при СД и к нарушению белкового состава плазмы крови. Это выражается
в уменьшении содержания альбуминов, повышении альфа-2, В- и Y-глобу-
линов. Нарушается обмен гликопротеидов, что проявляется в повышении
в сыворотке крови альфа-2-гликопротеидов, а также гексод, связанных с
белками. Нарушение обмена гликопротеидов обусловлено, с одной сторо-ны, дефицитом инсулина, а с другой - нарушением функции гипофиза, над-почечников и половых желез.
В процессе превращения белка в углеводы образуется аммиак, моче-
вина и другие продукты распада. В связи с этим при не леченном или де-компенсированном СД возникает гиперазотемы с последующей гиперазо-турией. Последняя обусловлена усиленным образованием аммиака как в
печени, так и в почках из глютамина.
2.2.1 ОПРЕДЕЛЕНИЕ ГЕМОГЛОБИНА.
Принцип: гемоглобин окисляют в метгемоглобин окисляют железосинеродистым калием (красная кровяная соль); образующийся с ацетонциангидрином окрашенный циан-метгемоглобин определяют как колориметрический.
Реактив: Трансформирующий раствор: ацетонциангидрин – 0,5 мг.; калий железосинеродистый – 0,2 г.; натрия гидрокорбанат – 1 г.; дистиллированная вода до 1 л. Раствор желтого цвета, прозрачный.
Калибровочный раствор гемоглобин цианида.
Специальное оборудование: фотоэлектроколориметр (ФЭК-56М).
Ход определения: В пробирку к 5 мл трансформирующего раствора добавляют 0,02 мл крови (разведение в 251 раз). Содержимое пробирки тщательно перемешивают и оставляют стоять 10 мин. Измеряют на ФЭКе при длине волны 500-560 нм (зелёный светофильтр) в кювете с толщиной слоя 1 см против холостой пробы (трансформирующий р-р.). Измеряют при тех же условиях в стандартный раствор.
Расчет содержания гемоглобина производят по калибровочному графику, построенному по стандартному раствору гемиглобинцианида, или по формуле:
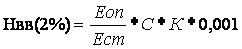 , где
, где
Еоп – экстинкция опытной пробы;
Ест – экстинкция стандартного раствора;
С – концентрация гемоглобинцианида в стандартном растворе, мг/%;
К – коэффициент разведения крови;
0,001 - коэффициент для пересчёта мг/100 мл. в г/100 мл
При использовании унифицированным гемоглобинцианидным методом нормальное содержание Нв у мужчин составляет от 132,0 – 164,0 г/л у женщин составляет от 115,0 – 145,0 г/л
2.2.2. Скорость оседания эритроцитов (унифицированный микрометод Панченкова).
Принцип: Смесь крови с цитратом при стоянии разделяется на два слоя (нижний - эритроциты, верхний – плазма). При этом СОЭ, т.е. величина столбика плазмы, бывает различной в зависимости от изменений физико – химических свойств крови.
Реактивы: 5% р-р трёхзамещённого цитрата натрия.
Специальное оборудование: Аппарат Панченкова, состоящий из штативов и капилляров. Пробирки и капилляры должны быть химически чистыми.
Ход определения: Перед использованием капилляра промыть цитратом натрия и заполнить им пробирку на ¼. Кровь набирают до метки "0". Устанавливают капилляр в штатив через час отмечают скорость оседания эритроцитов по высоте отстоявшегося слоя плазмы в мм.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА:
1. Фелик Ф., Бакстер Дж.Д., Бродус А.Е., Фромен Л.А. Эндокринология и
метаболизм: пер. с анг. - М.: Медицина, 1985, стр.7-212.
2. Балаболкин М.И. Сахарный диабет. - М.: Медицина, 1994, стр.7-37;
45-95.
3. Клиническая эндокринология: Руководство/под ред. Старковой Н.Т. -
М.: Медицина, 1991, стр.188-245.
4. Агеев А.К. Клеточный состав островков поджелудочной железы при ин-
сулинонезависимых формах сахарного диабета. - Клиническая медици-
на., 1984, т.62 № 8 стр. 93-98.
5. Алексеев Ю.П., Мирходжаев А.Х. Характер изменения секреции глю-
кагона у больных сахарным диабетом. - Проблема эндокринологии,
1978, т.24 № 4 стр.3-9.
6. Кило Ч., Уильямсон Дж., Ричмонд Д. Что такое диабет? Факты и реко-
мендации: пер. с англ. - М.: Мир, 1993, стр.18-20.
7. Потемкин В.В. Эндокринология: М.: Медицина, 1978, стр.202-287.
8. Кахновский И.М., Кузнецов Д.А., Давиденков Н.В. - Терапевтический