Классификация и жизненные циклы диатомовых
Страница 3
Вегетативное деление.
Основной способ размножения диатомовых водорослей – вегетативное деление клетки надвое. Деление обычно происходит ночью и на рассвете и осуществляется по-разному у разных видов, а также у одних и тех же видов в зависимости от условий среды. Наиболее интенсивного развития диатомовые водоросли достигают весной и осенью.
Перед делением в протопласте скапливаются капли масла, он увеличивается в размерах, раздвигает эпитеку и гипотеку так, что они соприкасаются лишь краями поясковых ободков. У многих видов митозу предшествует деление хлоропласта.
Многократные вегетативные деления приводят к постепенному уменьшению размеров клеток, получающих гипотеку материнской клетки. У некоторых видов они уменьшаются в 3 раз по сравнению с первоначальными.
Образование ауксоспор.
Восстановление первоначальных размеров клеток происходит во время прорастания покоящихся спор, покоящихся клеток и в результате полового процесса, сопровождающегося образованием ауксоспор. Считается, что до начала спорообразования клетки проходят внутреннюю перестройку, направленную на ликвидацию отклонений в метаболизме, вызванных нарушением соотношения объемов ядра и цитоплазмы в результате митотических делений. При этом происходит дифференциация клеток по их роли в репродуктивном процессе, так как не все клетки, достигшие определенного размера, переходят к спорообразованию.
У пеннатных диатомей половой процесс во всех случаях состоит в сближении двух клеток, в каждой из которых створки раздвигаются и происходит редукционное деление ядра, после чего гаплоидные ядра попарно сливаются и образуется одна или две ауксоспоры.
У центрических диатомей попарное сближение клеток отсутствует, и ауксоспора образуется из одной клетки, в которой сначала происходит деление материнского диплоидного ядра на четыре гаплоидных, два из них затем редуцируются, а два сливаются в одно диплоидное ядро и образуется ауксоспора.
Все диатомовые водоросли – диплоидные организмы, а гаплоидная фаза у них бывает только перед слиянием ядер в ауксоспоре. После оплодотворения образуется зигота, которая без стадии покоя начинает расти, увеличивается в размерах и превращается в ауксоспору («растущую спору»). При созревании ауксоспора превращается в инициальную клетку, значительно превосходящую по размерам исходную материнскую и иногда отличную от нее по форме. По положению относительно материнской клетки и связи с ней различают свободные, боковые, конечные, интеркалярные и полуинтеркалярные ауксоспоры. Диатомовые водоросли – единственная группа растительных организмов, в жизненном цикле которых есть стадия ауксоспорообразования.
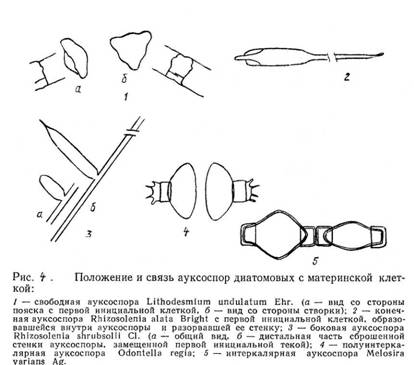
Половой процесс.
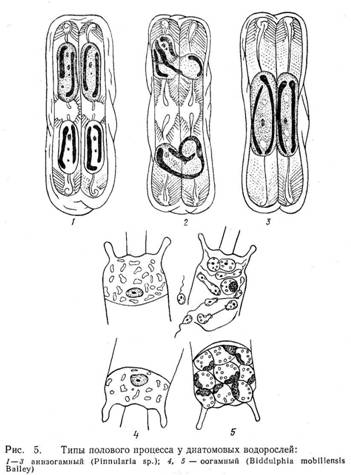
У диатомовых выявлено несколько типов полового процесса.
1. При изогамном половом процессе в двух материнских клетках образуется по две неподвижные гаметы, которые копулируют (сливаются) попарно (виды родов Amphora Ehr., Epithemia Breb., Rhopalodia O. Mull., Surirella Turp.).
2. Анизогамный (гетерогамный) половой процесс протекает двояко. В первом случае в ходе последовательных мейотического и митотического делений в каждой материнской клетке образуется по одной подвижной и одной неподвижной гамете. Подвижные гаметы передвигаются к неподвижным и сливаются с ними. Этот тип характерен дл большинства представителей семества Naviculaceae и некоторых видов рода Nitzschia. Во втором случае в одной клетке обе гаметы неподвижные, в другой – обе подвижные, переходящие в клетку с неподвижными гаметами. Такой тип анизогамии характерен для Nfvicula halophila (Grun.) Cl. и Synedra ulna (Nitzsch.) Ehr.
3. При оогамном половом процессе женская репродуктивная клетка (оогоний) производит одну яйцеклетку (виды рода Stephanopyxis Thr., Melosira varians Ag.) или две (Biddulphia mobiliensis Bail.), а мужская репродуктивная клетка (сперматогоний) образует два (Melosira varians Ag.) или четыре (Biddulphia rhombus, Cyclotella sp.) сперматозоида, оплодотворяющих яйцеклетку. У центрических диатомей, в отличие от других водорослей, сначала образуется большое число мелких сперматогониев, а мейоз происходит в самый последний момент, непосредственно перед обособлением гамет. Обычно же при гаметической или спорической редукции у других водорослей сначала совершается мейоз, затем при митотических делениях увеличивается число ядер и лишь после этого формируется большое число гамет или гаплоидных зооспор.
4. Автогамия - особый тип полового процесса, распространенный у части диатомовых. Заключается он в том, что ядро клетки предварительно делится с мейозом на 4 ядра, два из них разрушаются, и оставшиеся два ядра сливаются, образуя вновь диплоидное ядро. Автогамия не сопровождается увеличение числа особей, а лишь их омоложением.
Клетки, связанные с половым процессом, имеют некоторые структурные отличия от обычных клеток. Так, в сперматозоидах Lithodesmium undulatum Ehr. отсутствуют диктиосомы, а хлоропласты более мелкие и упрощенные; в аксонеме жгутиков сперматозоидов Lithodesmium undulatum Ehr. и Pleurosira laevis (Ehr.) Compere нет двух внутренних микротрубочек, жгутики покрыты мастигонемами – волосками диаметром 11 нм.
Микроспоры и покоящиеся споры.
У многих морских планктонных диатомей в клетках возникают микроспоры – мелкие тельца, образующиеся в количестве от 8 до 16 и более, у некоторых видов их бывает и более 100. Наблюдались микроспоры со жгутиками и без жгутиков, с хлоропластами и бесцветные. Образование микроспор особенно характерно для видов рода хетоцерос (Chaetoceros), у которых наблюдалось и их прорастание.
При неблагоприятных условиях диатомовые водоросли переходят в состояние покоя. В ходе образования покоящихся клеток протопласт передвигается к одному из концов клетки и, вследствие потери клеточного сока, сильно сжимается. Жизнедеятельность этих клеток возобновляется при наступлении благоприятных условий. Некоторые пресноводные планктонные озерные виды в зимний период погружаются на дно водоемов, где пребывают в состоянии покоя и пониженной жизнедеятельности до начала вегетационного периода.
У большинства диатомей образованию покоящихся спор предшествует деление вегетативной клетки на две, каждая из которых в дальнейшем становится материнской клеткой споры. Протопласт материнской клетки сжимается, округляется, на поверхности его возникает первичная створка споры, затем вторичная, которая выдвигается своими краями в края первичной. Содержимое споры гомогенно. Структура створок споры постоянна для каждого вида и отличается от структуры вегетативной клетки. Это один из наиболее важных видовых признаков. Как правило, материнская клетка производит одну экзогенную, полуэндогенную или эндогенную спору (у Rhizosolenia setigera Braight. – две). Зрелая экзогенная спора находится вне материнской клетки; одна створка зрелой полуэндогенной покоящейся споры включена в материнскую клетку, другая остается свободной; зрелая эндогенная покоящаяся спора находится внутри материнской клетки. Спора прорастает в вегетативную клетку, размер которой значительно превышает размер самой споры
Покоящиеся споры обычно образуют многие морские неритовые диатомеи, а также некоторые пресноводные виды. У представителей многих родов они возникают периодически как обычное явление в жизненном цикле.

Список используемой литературы:
1. Жизнь растений в шести томах./ Курсанов, А.Л.; Тахтаджян, А.Л.; Цицин, Н.В. и др.; гл. редактор Федоров, А.А – М.: Просвещение, 1977.;
2. Водоросли: справ./ Вассер, С.П.; Кондратьева, Н.В.; Масюк, Н.П. и др. – Киев: Наукова думка, 1989. – (Академия наук Украинской ССР институт ботаники им. Н.Г.Холодного).