Обмен веществ и энергии в живых организмах
Страница 2
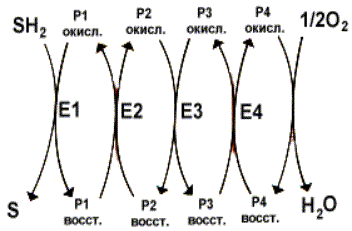
Перенос электронов и протонов с участием промежуточных переносчиков. SH2 - исходный донор протонов и электронов; P - промежуточные переносчики; E1, E2, E3, E4 - ферменты окислительно-восстановительных реакций
Промежуточными переносчиками в дыхательной цепи у высших организмов являются коферменты: NAD+ (никотинамид-адениндинуклеотид), FAD и FMN (флавинадениндинуклеотид и флавинмононуклеотид), кофермент Q (CoQ), семейство гемсодержащих белков - цитохромов (обозначаемых как цитохромы b, С1, С, А, А3) и белки, содержащие негеминовое железо. Все участники этой цепи разделены на четыре окислительно-восстановительные системы, связанные убихиноном (CoQ) и цитохромом С. Процесс начинается с переноса протонов и электронов от окисляемого субстрата на коферменты NAD+ или FAD. Это определяется тем, является ли дегидрогеназа, катализирующая первую стадию, NAD - зависимой или FAD - зависимой. Если процесс начинается с NAD+ , то следующим переносчиком будет FMN.
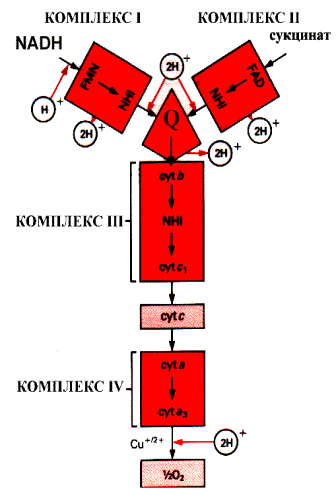
Последовательность промежуточных переносчиков протонов и электронов в дыхательной цепи
Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы ни был исходный субстрат, электроны и протоны от флавинов переносятся к коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками. NAD - зависимая дегидрогеназа катализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD+ является коферментом и выполняет роль акцептора водорода:
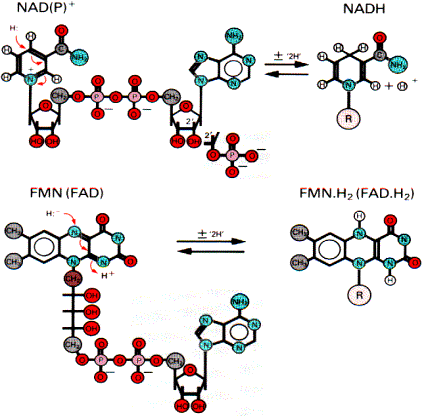
Коферменты дегидрогеназ
Символ 2Н+ означает два электрона и два протона, обычно переносимые в виде гидрид иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода». FAD - зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Коферментом является FAD, который является акцептором водорода от субстрата. NADH - дегидрогеназа катализирует окисление NADH и восстановление убихинона (CoQ). Переносчиком водорода является кофермент - FMN (комплекс 1). В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем передается на убихинон. Флавиновые коферменты (FAD и FMN) прочно связаны с ферментом как простетические группы, поэтому ферменты, в состав которых они входят, называются флавопротеины. Флавинмононуклеотид (FMN), или рибофлавин фосфат, неразрывно связан с белковой частью фермента. Строго говоря, FMN не является нуклеотидом, так как флавиновая часть связана с рибитолом, а не с рибозой.
Убихинон (кофермент Q) - производное изопрена:
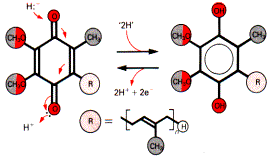
Название «убихинон» возникло из-за его повсеместной распространенности в природе. Кофермент Q действует как переносчик электронов на цитохромы.
Цитохромы - это гемопротеины - белки, содержащие в качестве прочно связанной простетической группы гем:
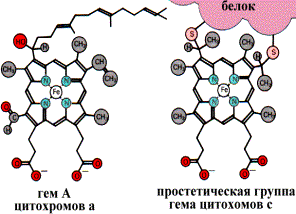
Простетическая группа гема в структуре цитохромов
Атом железа в геме может менять валентность, присоединяя или отдавая электроны:
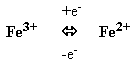
В дыхательной цепи цитохромы служат переносчиками электронов и располагаются соответственно величине окислительно-восстановительного потенциала следующим образом: B, С1, С, а, а3. Гемовые группы цитохромов связаны с белковой частью донорно-акцепторными связями между ионом железа и соответствующими аминокислотными остатками:
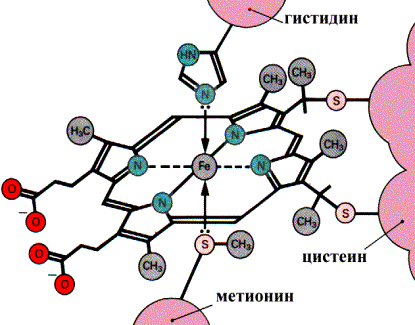
Связывание гема с белковой частью цитохрома С
В цитохромах С и С1 дополнительные ковалентные связи формируются между тиогруппами цистеина и боковыми винильными группами гема. QН2-дегидрогеназа (комплекс III) представляет собой комплекс цитохромов b и С1. Этот фермент катализирует окисление восстановленного кофермента Q и перенос электронов на цитохром С. Электроны последовательно переносятся атомами железа цитохромов b и С1, а затем поступают на цитохром С. Протоны после окисления QH2 освобождаются в раствор.
Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV). Цитохромоксидаза кроме гема содержит ионы меди, которые способны менять валентность и таким способом участвовать в переносе электронов:
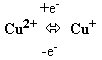
Цитохромоксидаза переносит электроны с цитохрома С на кислород. В переносе электронов участвуют сначала ионы железа цитохромов а и а3, а затем ион меди цитохрома а3. Молекула кислорода связывается с железом в геме цитохрома а3. Следовательно, переход электронов на кислород с иона меди цитохрома а3, происходит на молекуле фермента. Каждый из атомов молекулы кислорода присоединяет по два электрона и протона, образуя при этом молекулу воды.
Белки, содержащие негеминовое железо. Некоторое количество атомов железа в митохондриях связано не в геме цитохромов, а образует комплексы с другими белками. Эти белки называют также железосерными, так как атомы железа связаны с атомами серы цистеиновых остатков. Белки, содержащие негеминовое железо, участвуют в переносе электронов на нескольких стадиях, однако, не совсем ясны их локализация и механизм действия.
Окислительное фосфорилирование
Энергия, образующаяся при прохождении потока электронов по дыхательной цепи, используется для сопряженного фосфорилирования ADP. Эти два процесса взаимозависимы: окисление не может протекать в отсутствии ADP. Соотношение окисления и фосфорилирования определяется коэффициентом P/O (количество моль фосфорилированного ADP на 1/2 моль кислорода) коэффициент Р/О называется коэффициентом окислительного фосфорилирования и зависит от точки вхождения восстановительных эквивалентов в цепь транспорта электронов. Например Р/О=3, для субстратов, окисляемых NAD - зависимой дегидрогеназой , так как в дыхательной цепи есть три участка, где перенос электронов сопряжен с синтезом АТР. Не все субстраты передают электроны и протоны на NAD, некоторые окисляются FAD - зависимыми дегидрогеназами, которые переносят протоны и электроны сразу на убихинон, минуя первый комплекс. В этом случае Р/О=2. В действительности коэффициент фосфорилирования всегда меньше теоретической величины, потому что часть энергии, высвобождающейся при транспорте электронов, расходуется не на синтез АТР, а для переноса веществ через митохондриальную мембрану.
В сутки человек потребляет в среднем 27 моль кислорода. Основное его количество (примерно 25 моль) используется в митохондриях в дыхательной цепи. Следовательно, ежесуточно синтезируется 125 моль ATP или 62 кг (при расчете использовали коэффициент Р/О=2,5, то есть среднее значение коэффициента фосфорилирования). Масса всей АТР, содержащейся в организме, составляет примерно 20-30 г. Следовательно, можно сделать вывод, что каждая молекула АТР за сутки 2500 раз проходит процесс гидролиза и синтеза, что и характеризует интенсивность обмена АТР.
Сопряжение работы дыхательной цепи с процессом синтеза АТР
Существование такого сопряжения доказывается тем, что можно ингибировать образование АТР, не нарушая процесса транспорта электронов. Это достигается добавлением химических веществ, названных разобщителями. После удаления разобщителей синтез АТР восстанавливается. Изучение механизма сопряжении дает ответ на основные вопросы:
- каким образом транспорт электронов служит источником энергии?
- как эта энергия передается в реакцию ADP + Pi a АТР?