Обмен веществ и энергии в живых организмах
Страница 3
Существует несколько гипотез, объясняющих механизм сопряжения. Одной из них является хемоосмотическая теория. Цепь транспорта электронов функционирует как протонная (Н+)помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство. Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи. Перенос протонов приводит к возникновению разности концентрации Н+ с двух сторон митохондриальной мембраны: более высокая концентрация будет снаружи и более низкая - внутри. Митохондрия в результате переходит в «энергизованное» состояние, так как возникает градиент концентрации Н+ и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности.
Электрохимический потенциал способен совершать «полезную» работу, он заставляет протоны двигаться в обратном направлении, но мембрана непроницаема для них кроме отдельных участков, называемых протонными каналами. Обратный перенос протонов в матрикс является экзоэргическим процессом, высвобождающаяся при этом энергия используется на фосфорилирование ADP. Эту реакцию катализирует фермент Н+-АТР-синтетаза, располагающаяся в области протонных каналов на внутренней поверхности внутренней мембраны.
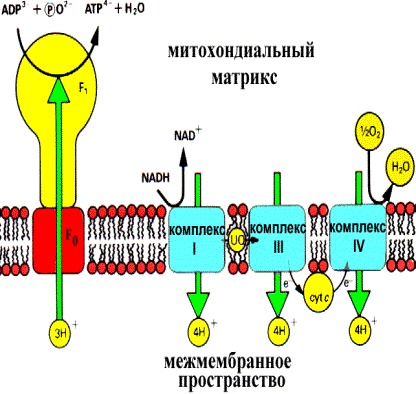
С опряжение цепи транспорта электронов и фосфорилирования ADP посредством протонного градиента
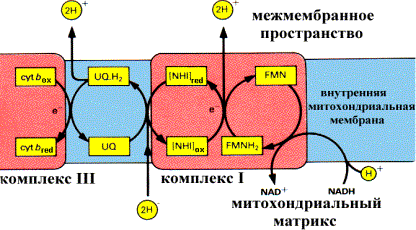
С труктура компонентов комплекса I, обеспечивающего функционирование «протонной помпы» при окислении NADH
Разобщение дыхания и фосфорилирования
Убедительные экспериментальные доказательства в пользу описанного механизма сопряжения дыхания и фосфорилирования были получены с помощью ионофоров. Молекулы этих веществ, как правило, липофильны и способны переносить ионы через мембрану. Например, 2,4-динитрофенол (протонофор) легко диффундирует через мембрану, в ионизированной и неионизированной форме, перенося протоны в сторону их меньшей концентрации в обход протонных каналов. Таким образом, 2,4-динитрофенол уничтожает электрохимический потенциал, и синтез АТР становится невозможным, хотя окисление субстратов при этом происходит. Энергия дыхательной цепи в этом случае полностью рассеивается в виде теплоты. Этим объясняется пирогенное действие разобщителей. Разобщающим действием обладают гормон щитовидной железы - тироксин, а также некоторые антибиотики, такие как валиномицин и грамицидин.
Дыхательный контроль
Скорость дыхания митохондрий может контролироваться концентрацией ADP. Это объясняется тем, что окисление и фосфорилирование жестко сопряжены. Энергия, необходимая клетке для совершения работы, поставляется за счет гидролиза АТР. Концентрация ADP при этом увеличивается; в результате создаются условия для ускорения дыхания, что и ведет к восполнению запасов АТР.
Ингибиторы цепи транспорта электронов и окислительного фосфорилирования
Ингибиторы, блокирующие дыхательную цепь, действуют в определенных местах, препятствуя работе дыхательных ферментов (KCN, барбитураты, ротенон). Существуют также вещества, ингибирующие окислительное фосфорилирование.