Генетика пола, наследование, сцепленное с полом
Страница 4
Кроссинговер может происходить не только во время мейоза, но и митоза, тогда его называют митотическим кроссинговером. Частота митотического кроссинговера значительно ниже мейотического. Тем не менее ег также можно использовать для генетического картирования.
Мейотический кроссинговер осуществляется после того, как гомологичные хромосомы в зиготенной стадии профазы I соединяются в пары, образуя биваленты. В профазе I каждая хромосома преджставлена двумя сестринскими хроматидами, и перекрест происходит между хроматидами.
 Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган допустил, что перекрест между хроматидами гомологичных хромосом может происходить одновременно в нескольких точках кроссинговер, происходящий лишь в одном месте, называют одиночным кроссинговером, в двух точках одновременно - двойным, в трех - тройным и т.д., т.е. кроссинговер может быть множественным.
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган допустил, что перекрест между хроматидами гомологичных хромосом может происходить одновременно в нескольких точках кроссинговер, происходящий лишь в одном месте, называют одиночным кроссинговером, в двух точках одновременно - двойным, в трех - тройным и т.д., т.е. кроссинговер может быть множественным.
Пусть, например, в гомологичной паре хромосом содержатся три пары аллелей в гетерозиготном состоянии
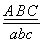
Тогда перекрест, произошедший только в участке между генами А и В или между В и С, будет одинарным. В результате одинарного перекреста возникают в каждом случае только две кроссоверные хромосомы
aBC и Abc или Abc и aBC.
Каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках. Таким образом, двойные кроссинговеры сокращают регистрируемое расстояние между генами.
Вместе с тем между обменами на соседних участках хромосом существует взаимовлияние, названное интерференцией. Такое взаимовлияние можно выразить количественно. Для этого составляют реально наблюдаемую частоту двойных кроссинговеров с частотой, теоретически ожидаемой на основе предположения о том, что обмены на соседних участках происходят независимо друг от друга. Степень и характер интерференции измеряется величиной коинциденции (С). Коинциденцию оценивают как частное от деления реально наблюдаемой частоты двойных кроссоверов на теоретически ожидаемую частоту двойных кроссоверов. Последнюю величину получают, перемножая частоты кроссинговера на соседних участках.
Величину интерференции (I) определяют по формуле I=1-C. Если С<1, то интерференция положительная, т.е. одинаковый обмен препятствует обмену на соседнем участке хромосомы. Если С>1, то интерференция отрицательная, т.е. один обмен как бы стимулирует дополнительные обмены на соседних участках. В действительности существует только положительная интерференция при реципрокной рекомбинации - кроссинговере, а кажущееся неслучайным совпадение двух и более обменов, характерное для очень коротких расстояний - результат нереципрокных событий при рекомбинации.
Таким образом, при карплеровании генов в группах сцепления на основе изучения частот рекомбинации необходимо учитывать две противоположные тенденции. Двойные обмены “сокращают” расстояния между генами, и интерференция препятствует множественным обменам, вероятность которых увеличивается с расстоянием.
В обобщенном виде зависимость частоты рекомбинации от реального расстояния с учетом множественных обменов описывает функция Дж. Холдэйна.
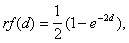
где rf - картирующая функция (в нашем случае - это частота учитываемых кроссинговеров), d - реальное расстояние, на котором происходят обмены, e - основание натурального логарифма.
При изучении множественных обменов и интерференции между ними используют тетрадный анализ. Для этого рассматривают тригибридное скрещивание (ABC x abc) по сцепленным генам. Учитывая, что кроссинговер происходит на стадии 4-х хроматид, возможны три типа двойных обменов. Это двойные двухроматидные обмены, двойные треххроматидные обмены и двойные четыреххроматидные обмены только между несестринскими хроматидами, последствия которых генетически различимы (рис. 4).
3.4. Группы сцепления и карты хромосом у человека.
|
 9 9
1. Lu Se
 R El R El
 2. 2.
10
 N I N I
3.

Рис. 7. Генетические карты аутосом человека. |
У человека 23 пары хромосом. Это указывает на наличие у него 23 групп сцеплений, для каждой из которых надо построить линейные карты взаиморасположения генов. Хорошо установлены группы сцепления, касающиеся трех пар аутосом. Одна группа сцепления несет в себе локус 1, где локализованы аллели групп АВО и локус, содержащий дефекты локтей и коленной чашечки (N). Расстояние между этими генами равно 10% кроссинговера. Вторая группа сцепления в аутосоме содержит локус Rh, где локализованы аллели резус-фактора, и локус эллиптоцитоза (El) доминантной мутации, вызывающей овальную форму эритроцитов. Расстояние между этими локусами равно 3%. Третья аутосома имеет в себе локусы группы крови Лютеран (Lu) и локус секреции (Se). Группы крове Лютеран содержат систему из двух аллелей Lua и Lub. Аллели - секреторы (se) обуславливают выделение в разных тканях организма, и, в частности в слюне, растворимых в воде антигенов АВО. Люди с рецессивными аллелями этого локуса (H) не выделяют водорастворимых антигенов. Действие аллеля касается групп крови с антигеном АВО и антигеном групп крови Лютеран. Расстояние между локусами Lu и Se равно 9%.
Четвертая генетическая карта касается Х-хромосомы (рис. 8).
|
 25 10 25 10
 n m c h n m c h
50
Рис. 8. Генетические карты Х-хромосо-мы человека. |
Начальный период в составлении карт хромосом человека очень знаменателен. Будущая медицина и антропология будут связаны с использованием этих данных. Для борьбы с врожденными болезнями и многими отрицательными биологическими сторонами человека раскрытие генетического строения его 23 пар групп сцепления с их точными линейными картами генов и знание тонкого строения отдельных генов сыграют величайшее значение.
4. Заключение.
Таким образом, генетика занимает важное место в жизни человека. Именно она объясняет механизмы наследования признаков человека, как патологических, так и положительных. Так, пол человека - это менделирующий признак, наследуемый по принципу обратного скрещивания.
У женщин пол гетерогаметен (XY), у мужчин гомогаметен. Среди признаков, подчиняющихся законам Г. Менделя, существуют признаки наследуемые сцепленно. Однако сцепление часто бывает неполным, причина тому кроссинговер, который имеет важное биологическое значение - лежит в основе комбинативной изменчивости.