Поведение чайки
Страница 2
В различные сроки после вылупления птенцам предъявляли модель головы взрослой птицы. Поведение птенцов регистрировалось на кинопленке, анализ которой позволял определять процент точных клевков. Во всех пяти группах точность с возрастом увеличивалась, но только в двух контрольных группах она достигала нормального уровня (свыше 75%). В отличие от зрительной депривации, оказывавшей заметное влияние на точность клевания, исключение нормальной активности, связанной с вылуплением, практически не влияло на результаты тестирования.
Наиболее осторожной интерпретацией этих результатов мог бы быть следующий вывод: зрительный опыт необходим для достижения максимальной точности клевания, но некоторое повышение точности происходит и в условиях зрительной депривации. В последнем случае улучшение может быть результатом совершенствования позных реакций. Здесь тоже возможно влияние индивидуального опыта, так как весьма вероятно, что позные реакции эффективно отрабатываются у птенцов в условиях темного инкубатора.
Как научается птенец выбирать правильное расстояние для точного клевка? Наблюдения над только что вылупившимися птенцами позволяют предположить, что здесь действует механизм саморегуляции, основанный на восприятии расстояния. Если неопытный птенец встанет слишком близко к модели, то удар оказывается настолько сильным, что он может отбросить птенца назад на 2-3 см. И наоборот, когда птенец находится слишком далеко от цели, он не достает до нее при клевке и падает вперед на 4-5 см. Старшие птенцы редко допускают такие серьезные ошибки; можно думать, что с возрастом они методом проб и ошибок научаются выбирать наилучшее расстояние.
Исследователи нередко рассматривали пищевую мотивацию (голод) как результат научения. Эксперименты позволяют предполагать, что по крайней мере в основе голода лежит нечто врожденное. Ряд опытов показал, что у накормленных досыта птенцов, как и следовало ожидать, частота клевков возрастает с увеличением интервала времени после кормления. Однако такая же картина наблюдалась и у птенцов, которым мы ни разу не доказали случая испытать чувство голода. Птенцов, вылупившихся в темном инкубаторе, искусственно кормили до насыщения на вторые сутки жизни, а затем тестировали на свету с моделями головы. Через час после кормления средняя частота клевков составляла у них 3,1 в минуту, а через два часа-5,1 (разница статистически достоверна).
Детали движений при клевке во время пищевой реакции анализировались с помощью высокоскоростной киносъемки. У птенца в возрасте нескольких дней пищевая реакция, к этому времени уже вполне сформированная, включает четыре основных компонента: 1) открывание и последующее закрывание клюва; 2) движение головы вверх и вперед по направлению к голове взрослой птицы, а затем вниз и назад; 3) поворот головы в сторону в расчете на то, чтобы захватить клювом клюв взрослой птицы, и обратный поворот головы; 4) легкий толчок ногами вверх и вперед (см. рис. 114). Покадровый анализ фильма позволил выявить значительную изменчивость временных отношений между этими компонентами, как у каждого отдельного птенца, так и у различных птенцов. С возрастом эта изменчивость несколько уменьшается, что, вероятно, связано с улучшением координации движений. Среди интересных деталей, выявленных при анализе кинолент, можно отметить тот факт, что у птенцов, выросших в естественных условиях, поворот головы в сторону наблюдался с возрастом все чаще. Чтобы выяснить, насколько это зависит от зрительного опыта, мы проанализировали фильмы, заснятые во время тестов на точность клевания во всех пяти группах птенцов. Оказалось, что у птенцов, не имевших опыта клевания, сколько-нибудь заметный поворот головы наблюдался редко. У птенцов, выросших в гнезде, в первые сутки жизни поворотов головы не бывает, но затем эта реакция появляется и быстро совершенствуется. Не известно, какова конкретная роль индивидуального опыта в таком развитии поведения, но снятые фильмы дают основание для некоторых предположений. Иногда, когда птенец клюет с раскрытым клювом, клюв взрослой птицы, расположенный не строго вертикально, может оказаться между челюстями птенца. Тогда движение головы птенца по инерции вперед поневоле приводит к ее повороту в сторону. Возможно, именно так птенец научается поворачивать голову, чтобы схватить клюв взрослой птицы.
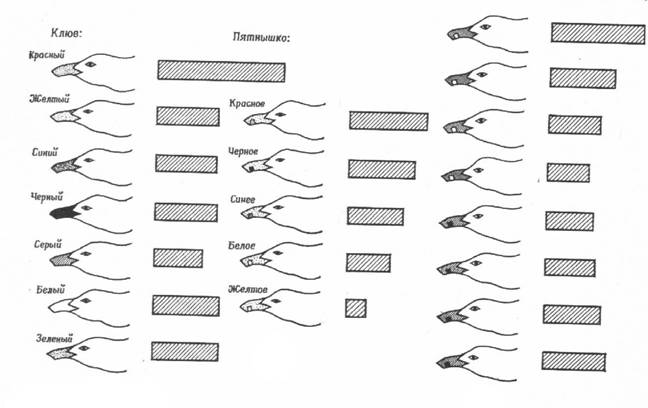
Один из самых интересных вопросов, связанных с пищевым поведением птенцов - это вопрос о том, как птенцы узнают своих родителей. Наблюдения за развитием птенцов в естественных условиях показали, что птенцы иногда клюют не только клюв взрослой птицы, но и иные предметы, в том числе другие части тела родителя. Правда, большинство клевков все же нацелено на клюв, особенно в более позднем возрасте. Это позволяет предположить, что недавно вылупившиеся птенцы представляют себе родителей довольно смутно, но с возрастом, по мере накопления опыта, образ взрослой птицы становится у них все более четким. Эту проблему исследовали с помощью набора различных моделей головы и клюва (на рисунке показаны модели и относительная частота ударов по ним клювом).
Изменяя или устраняя отдельные части таких моделей, смогли выявить наиболее эффективные стимулы для пищевой реакции. Обычно модель закреплялась на стержне, который мог колебаться в такт с метрономом, так что скорость движения модели была всегда известна. В каждом опыте птенцу в случайном порядке предъявлялись пять моделей и при этом регистрировалось число клевков за определенное время (обычно за 30 секунд). Задача состояла в том, чтобы выявить наиболее эффективные стимулы для птенцов, находившихся в темноте в течение суток с момента вылупления до начала эксперимента. В первых трех опытах птенцы одинаково реагировали на все модели, кроме той, у которой не было клюва. Таким образом, показано, что новорожденные птенцы реагируют в основном на клюв родителей, а не на особенности головы (даже само наличие головы для птенцов этого возраста несущественно).
Хотя оптимальный стимул для реакции клевания, очевидно, несложен, он, видимо, должен обладать какими-то чертами, отличающими его от прочих окружающих объектов, таких как красные лапы родителей или травинки, поскольку клевки птенцов редко бывают
нацелены на посторонние предметы. Этот вопрос исследовали с помощью деревянных палочек, окрашенных в красный цвет. Палочку предъявляли птенцу в горизонтальном или вертикальном положении, причем в обоих случаях она либо была неподвижна, либо перемещалась в горизонтальном или вертикальном направлении. Любая вертикальная палочка вызывала клевание с большей частотой, чем любая горизонтальная, а из вертикальных наиболее эффективной оказалась палочка, движущаяся в горизонтальном направлении. Это хорошо согласуется с естественной ситуацией, когда вертикально
 расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
Дальнейший анализ результатов показал, что вертикально движущийся вертикальный стимул вызывал не больше реакций, чем неподвижный вертикальный. Кроме того, и горизонтальное, и вертикальное перемещение горизонтального стимула было одинаково действенно и, как это ни странно, превосходило по эффективности неподвижный горизонтальный стимул. Наиболее осторожная интерпретация этих результатов может состоять в том, что адекватными стимулами для пищевой реакции птенцов служат два типа движения: 1) горизонтальное перемещение и 2) движение в направлении, перпендикулярном продольной оси стимула. Это позволяет объяснить, почему неподвижная вертикальная палочка оказывается не менее эффективным стимулом, чем вертикальная палочка, движущаяся вертикально. Дело в том, что в обоих случаях отсутствуют движения указанных типов. Однако в случае горизонтальной палочки вертикальное движение перпендикулярно продольной оси стимула и потому так же эффективно, как горизонтальное, и реакция здесь при обоих движениях более интенсивна, чем при полной неподвижности стимула.
На следующем этапе мы испытывали вертикальные стимулы различных диаметров, и пять скоростей движения. Палочка диаметром около 8 мм была наиболее эффективна вне зависимости от скорости движения. Скоростью, вызывающей наиболее интенсивную реакцию независимо от диаметра палочки, оказалась скорость 12 см/с. Эти результаты показывают, насколько точно реактивность в процессе эволюции подгоняется к параметрам стимулов. Толщина клюва взрослой птицы в вертикальном направлении 10,6 мм, а в горизонтальном-3,1 мм, так что средняя толщина составляет как раз около 8 мм. Кроме того, как показал анализ фильмов, снятых в естественных условиях, средняя скорость движения клюва при кормлении равна около 14,5 см/с.