Бiохiмiя трансгенноi картоплi в умовах Украiни
МРЖНРЖСТЕРСТВО АГРАРНОРЗ ПОЛРЖТИКИ УКРАРЗНИ
СУМСЬКИЙ ДЕРЖАВНИЙ АГРАРНИЙ УНРЖВЕРСИТЕТ
АГРОНОМРЖЧНИЙ ФАКУЛЬТЕТ
Кафедра захисту рослин
Завiдувач кафедрою_____________________
А.К.Мiшньов
ДИПЛОМНА РОБОТА
на тему: ВлБiохiмiя трансгенноi картоплi в умовах
УкраiниВ»
Для одержання квалiфiкацii спецiалiста
за спецiальнiстю 7.130105 тАУ Захист рослинВ»
Виконавець _________________ (ЗагорулькоО.)
Науковий керiвник ______________ (Чiванов В.Д.)
Консультант з екологiчноi
експертизи ________________ (Жатова Г.О.)
Консультант з питань
технiки безпеки_________________ (Шандиба О.Б.)
Суми - 2001
ЗМРЖСТ
Стор.
Вступ.........................................................
1. Огляд лiтературитАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж.
1.1. Загальна характеристика фiзико-хiмiчних властивостей глiкоалкалоiдiв
рослин родини Solanaceae................тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж
1.1.1. Глiкоалкалоiди рослин картоплi та iх будоватАжтАжтАжтАжтАжтАжтАжтАжтАжтАж.
1.1.2. Глiкоалкалоiди як токсичнi сполукитАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж
1.1.3. Роль глiкоалкалоiдiв в явищi iмунiтета до бактерiальних, грибкових захворювань i шкiдникiв та iх значення в селекцii нових сортiв
картоплiтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж
2. Природнi умови господарства...................................
3. Методика проведення дослiджень...............................
4. Результати дослiджень.........................................
5. Охорона працi................................................
6. Охорона навколишнього середовища............................
Висновки.....................................................
Список використаноi лiтератури................................
ВСТУП
Пiдвищений тАЬтисктАЭ на органiзм людини ксенобiотичних сполук природного та штучного походження обумовлюi необхiднiсть поглибленого вивчення таких сполук, i в першу тих iз них, котрi входять до складу найбiльш поширених харчових продуктiв, зокрема картоплi. Глiкоалкалоiди картоплi являють собою групу вторинних метаболiтiв, яким притаманнi найрiзномантнiшi види бiологiчноi активностi [ ]. Так, дослiдами in vitro доведена здатнiсть цих сполук спричинювати хромосомнi аберацii, порушувати цiлiснiсть лiпосом та природнiх бiомембран тощо [ ]. Останнi свiдчить на користь того, що глiкоалкалоiдам притаманна потенцiйна мутагенна i, можливо, канцерогенна активнiсть [ ]. Виходячи з наведеного, цiлком природньою i пiдвищена зацiкавленiсть в дослiдженнi глiкоалкалоiдiв з боку фахiвцiв в галузях медичноi токсикологii i генетики людини, тим бiльше, що в останнi роки на свiтовий ринок виходять новi сорти продовольчоi картоплi, отриманоi як шляхом тАЬкласичноiтАЭ селекцii за участi дикоростучих видiв роду Solanum [ ], так i за допомогою генетичноi iнженерii (New LeafÒ, Monsanto, США). Обидва пiдходи не виключають бiосинтезу в тканинах рослин картоплi, зокрема бульбах, окрiм характерних для культурноi картоплi глiкоалкалоiдiв a-Соланiну та a-Чаконiну тАЬмiнорнихтАЭ глiкоалкалоiдiв (солакаулiн, соласолiн, соламаргiн, демiссiн, томатiн, лептiн РЖ i РЖРЖ, лептiнiн-РЖ i РЖРЖ тощо), якi ведуть походження з дикоростучих попередникiв, або синтезуються de novo [ ]. Останнiм притаманнi токсикологiчнi характеристики, вiдмiннi вiд добре вiдомих аналогiчних показникiв a-Соланiну та a-Чаконiну. Якщо прийняти до уваги встановлений факт щодо синергетичного багаторазового пiдсилення бiологiчноi активностi сумiшi двох рiзних за хiмiчною структурою глiкоалкалоiдiв [ ], то стаi цiлком обгрунтованою необхiднiсть дослiджень кiлькiсних та якiсних показникiв глiкоалкалоiдiв продовольчоi картоплi паралельно з селекцiiю та молекулярною iнженерiiю, успiшнiсть яких залежить вiд наявностi нових високочутливих методiв фiзико-хiмiчного аналiзу. Складнiсть аналiзу сумарноi фракцii глiкоалкалоiдiв з рослин картоплi повтАЩязана, зокрема з тим, що субфракцiя тАЬмiнорнихтАЭ глiкоалкалоiдiв не перевищуi за масою 0,5-5% [ ].
Останнiм часом в аналiзi бiомолекул набули широкого розповсюдження методи мтАЩякоiонiзацiйноi мас-спектрометрii, зокрема плазмено-десорбцiйна мас-спектрометрiя (PDMS). Зважаючи на це, нами зроблена спроба проаналiзувати фракцiю глiкоалкалоiдiв бульб та квiтiв картоплi трансгенних сортiв NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic за допомогою PDMS мас-спектрометрii. Таким чином, метою наших дослiджень був докладний аналiз глiкоалкалоiдiв бульб та квiтiв картоплi трансгенних сортiв NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic новiтнiм мас-спектрометричним методам.
ОГЛЯД ЛРЖТЕРАТУРИ
1.1. Загальна характеристика фiзико-хiмiчних властивостей глiкоалкалоiдiв рослин родини Solanaceaе
Рослини картоплi культурних та диких видiв мiстять серед численних органiчних сполук групи вторинних метаболiтiв глiкоалкалоiди [ ], найважливiшими з яких i a-соланiн, a-чаконiн, демiссин, лептiни I,II,III, соламарiн, коммерсонiн [ ]. Фiзiологiчнi функцii глiкоалкалоiдiв пов'язанi з явищем стiйкостi рослин картоплi проти фiтопатогенiв i шкiдникiв [ ]. Крiм того, глiкоалкалоiди беруть безпосередню участь в бiохiмiчних процесах, якi притаманнi тканинам листкiв, пагонiв, квiток, бульб рослин картоплi (цвiтiння, видова сумiснiсть, утворення насiння); обумовлюють разом з iншими вторинними метаболiтами смаковi якостi та придатнiсть картоплi для переробки [ ], а також мають певне значення як токсичнi, ембрiотропнi i канцерогеннi сполуки природнього походження [ ]. Зважаючи на це, багато перспективних за врожайнiстю та iншими господарсько-корисними якостями сортiв картоплi не були рекомендованi для широкого використання саме завдяки пiдвищеному вмiсту глiкоалкалоiдiв [ ]. На думку вiтчизняних та зарубiжних [ ] дослiдникiв, кiлькiснi та якiснi характеристики глiкоалкалоiдiв нових сортiв i гiбридiв картоплi заслуговують на поглиблене вивчення. Одним з найважливiших питань, повтАЩязаних з бiохiмiiю та фiзiологiiю глiкоалкалоiдiв, яке i досi не знайшло однозначного вирiшення, i встановлення корелятивних звтАЩязкiв мiж параметрами метаболiзму глiкоалкалоiдiв та ступенем стiйкостi сортiв картоплi проти шкiдникiв i фiтопатогенiв [ ]. РЖншим дискусiйним питанням i невизначенiсть звтАЩязкiв мiж кiлькiсним накопиченням глiкоалкалоiдiв в тканинах рослин картоплi рiзних сортiв, зокрема бульбах i умовами вирощування (погодно-клiматичнi умови протягом конкретного вегетацiйного сезону, тип грунтiв, удобрення тощо), а також умовами зберiгання (температура, освiтленiсть, вологiсть тощо)[ ]. На думку провiдних вчених дискусiйнi та суперечливi результати в цьому напрямку дослiдження отриманi тому, що, по-перше, на iнтенсивнiсть бiосинтезу вторинних метаболiтiв взагалi i глiкоалкалоiдiв зокрема, рiзноспрямовано впливаi велика кiлькiсть факторiв зовнiшнього середовища i це, разом з пiдвищеною лабiльнiстю та чутливiстю ферментних систем метаболiзму глiкоалкалоiдiв до зазначених факторiв обумовлюi суттiвi розбiжностi, про якi згадувалось вище; i, по-друге, бiльшiсть раннiх робiт були проведенi без урахування рiзноманiтностi глiкоалкалоiдiв за хiмiчною структурою тАУ в межах цих дослiджень визначали тiльки iх сумарну кiлькiсть ваговим або колориметричним методами [ ].
Проте, глiкоалкалоiди картоплi являють собою досить велику групу сполук, до складу якоi входять ВлмажорнiВ» (a-соланiн та a-чаконiн у культурних видiв картоплi) i численнi ВлмiнорнiВ» (b-соламарiн, демiссидiн, 5b-соланiдан-3a-ол, томатiденол) компоненти. Крiм того, тканини рослин картоплi мiстять спецiалiзованi ферментнi системи, якi модифiкують iнтактнi молекули глiкоалкалоiдiв з утворенням численних промiжних сполук, фiзiологiчна роль яких вичерпно не зтАЩясована (b-соланiн, b-чаконiн, g-соланiн, g-чаконiн тощо)[ ]. Таким чином, дослiдження статичних i динамiчних кiлькiсних та якiсних параметрiв глiкоалкалоiдiв рослин картоплi рiзних сортiв в процесах вирощування в умовах конкретних клiматичних зон, зберiгання та переробки, а також пiд впливом фiтопатогенiв, мають певнi перспективи за умов використання з метою аналiзу цих сполук нових високоiнформативних фiзико-хiмiчних методiв дослiджень [ ]. В Сумському ДАУ та РЖнститутi прикладноi фiзики НАНУ (м.Суми) в 1994-2000 рр. в межах проекту ВлМтАЩякоiонiзацiйна мас-спектрометрiя в агробiологiiВ» провели цикл дослiджень глiкоалкалоiдiв рослин картоплi за допомогою новiтнього фiзико-хiмiчного методу аналiзу полярних, термолабiльних, нелетючих органiчних сполук тАУ плазмено-десорбцiйноi часопрольотноi плазмено-десорбцiйноi мас-спектрометрii з iонiзацiiю уламками подiлу 252Cf (МС-ПДМС)[ ].
Алкалоiди являють собой велику групу рослинних речовин рiзного хiмiчного складу, яким притаманна специфiчна дiя на органiзм ссавцiв та людини. Алкалоiди i азотовмiськими гетероциклiчними сполуками, що мiстять в своiму складi атом азоту. Загальною властивостю даноi групи речовин i iх надзвичайно сильна фiзиологiчна дiя на органiзм тварин та людей. Бiльшiсть алкалоiдiв дiють на нервову систему: в невеликих дозах здiйснюють збуджуючий вплив, в великих - пригнiчують, а в ще бiльших i сильнодiючими отрутами [ ]. Фiзiологiчне значення алкалоiдiв в життiдiяльностi самих рослин досi не з`ясовано. До цього часу не вдалося ще з великою достовiрностю показати мiсце алкалоiдiв в життiво важливих процесах обмiну речовин, з iншого боку немаi неспростовних доказiв того, що алкалоiдоноснi види рослин можуть iснувати без алкалоiдiв [ ].
В рядi робiт вiдзначаiться вплив алкалоiдiв на обмiн речовин у рослин, виникнення квiтiв, формування органел та органiв рослин, рiст клiтин, подiл ядер i т.д. Було доведено, що алкалоiди в клiтинах рослин якимось чином iзольованi вiд внутрiшнього середовища клiтини. Так, розчини нiкотину, нанесенi на рослиннi тканини табаку, чинять всебiчну негативну фiтотоксичну дiю навiть при концентрацii, рiвнiй iх концентрацii в тканинах. Доведено, що високий вмiст алкалоiдiв часто пов`язаний зi зниженням сили росту рослин. РЖнакше кажучи, у рослин в перiод вегетацii та спокою iснуi тiсний взаiмозв`язок: Влбiлковий азот - алкалоiдиВ». Цими причинами, на думку Горяiва, пояснюiться змiна складу алкалоiдiв, а в деяких випадках i iх повна вiдсутнiсть в тканинах алкалоiдних рослин (Solsola richteri. Girgensonia Oppositiflora. Anabasis turkestanika) [ ].
1.1. 1. Глiкоалкалоiди рослин картоплi та iх будова.
Глiкоалкалоiди, якi мiстяться в бульбах картоплi, ранiше називали загальним термiном ВлсоланiнВ». В наш час встановлено, що так званий ВлсоланiнВ» являi собою сумiш якнайменше 6 рiзних глiкозiдних сполук: a-, b-, g-соланiна та a-, b-, g-чаконiна. Цi сполуки мають один i той же аглiкон (Влкорове ядроВ») тАУ соланiдин i вiдрiзняються мiж собою складом глiкозидного ланцюжка (Рис.1). Трисахарiди - a-соланiн i a-чаконiн i основними формами i складають в тканинах картоплi бiля 95% усiii суми глiкоалкалоiдiв групи соланiна [ ]. Крiм a-, b-, g-соланiна та a-, b-, g-чаконiна, вiдомi iншi глiкоалкалоiди пасльонових: солакаулiн, соласолiн, соламаргiн, демiсiн, томатiн, лептiн, лептiнiн-1, лептiнiн-2 та iншi. Всi гликоалкалоiди складаються з двох основних складових, виникаючих при кислотному гiдролiзi цих речовин тАУ аглiкона, який i i власне ВлалкалоiдомВ», i декiлькох моносахарiв, якi знаходяться в молекулi глiкоалкалоiда в виглядi глiкозiдного ланцюга, зв`язаного з аглiконом [ ]. Глiкоалкалоiди культурноi та дiкоi картоплi (Solanum) можна систематизувати в окремi групи. До групи соланiдiна вiдносяться: a-, b-, g-соланiн та a-, b-, g-чаконiн, солакаулiн i глiкоалкалоiд, видiлений з групи Akaulia. РЖншу групу алкалоiдiв складають сполуки, аглiконом яких i соласодiн. Сюди входять соласонiн i соламаргiн. Соласонiн i соламаргiн i основними алкалоiдами Solanum nigrum, Solanum laciniatum бiльшостi iнших видiв Solanum
[ ].
Соласодамiн знайдений в листках Solanum auriculatum i Solanum magrinatum. В групу, яка мiстить в якостi аглiкона демiсiдiн, входить демiсин, знайдений у Solanum demissum i глiкоалкалоiд, видiлений iз Solanum polyadenium. Лептiни тАУ глiкоалкалоiди,
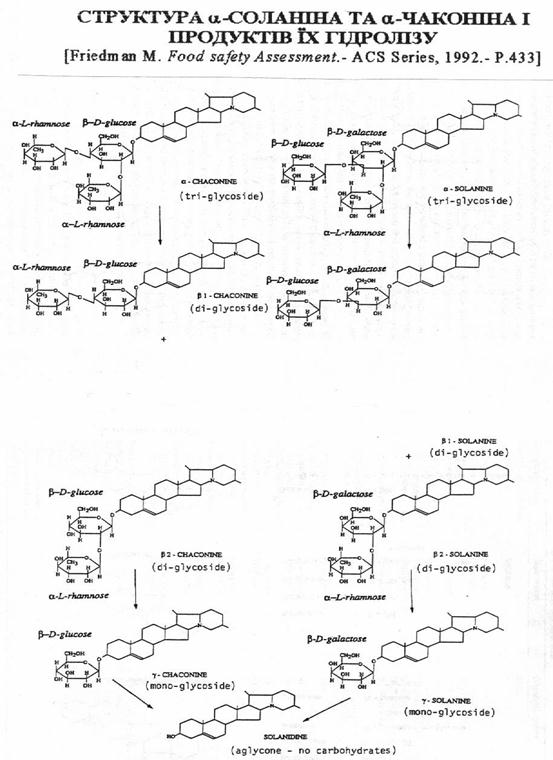
Рис.1. Хiмiчна структура аглiконiв, знайдених в рослинах картоплi [ ].
знайденi Куном i Левом, виявленi у Solanum chacoense. Групи Eutuberosa, Andigena, i Commersonia мiстять тiльки - соланiн, цi види мiстять також чаконiн. Цi два глiкоалкалоiди зустрiчаються завжди разом, що доведено хроматографiчними дослiдженнями. Демiсин присутнiй в рослинах групи Demissa i Acaulia, за виключенням Solanum acaulia, в складi якого знайдено солакаулiн. Демiсин знайдений також у Solanum jamesii. У форм, якi не дають бульб, крiм a-соланiна, який i тут частiше всього зустрiчаiться, найбiльшу групу складають форми, якi мiстять томатiн. РЖ тiльки iдина форма Solanum poliadenium, яка створюi бульби, мiстить томатiн. РЖншi алкалоiди зустрiчаються тiльки в деяких форм, нприклад, соласодамiн тАУ у Solanum sodomaneum i Slanum aviculare.
Всi алкалоiди картоплi належать до стероiдноi групи похiдних циклопентанопергiдрофенантрена i складаються з алкалоiда (аглiкона) та цукру, звтАЩязаного з ним глiкозiдним звтАЩязком. Аглiкон маi стероiдну будову, мiстить атоми азоту, який утворюi третинну або вторинну амiногрупу. Прикладом першого типу являiться соланiдiн, другого-томатiдiн i соласодiн. Глiкозiдна частина являi собою моно-, дi-, три-, i тетрасахарiди; до iх складу може входити глюкоза (Glu), галактоза (Gal), рамноза (Rha) i ксiлоза (Xyl). Наприклад, a-соланiн складаiться з аглiкона-соланiдiна i трисахарiда, який при гiдролiзi кислотою розпадаiться на галактозу, глюкозу i рамнозу. Соланiн маi розгалужений сахарний ланцюжок: безпосередньо з аглiконом зв`язана галактоза, а глюкоза i рамноза складають боковi ланцюги. Подiбну будову мають глiкозiднi частини глiкоалкалоiдiв демiсiну, томатiну, a-чаконiну, соласодiну i соламаргiну [ ].
Пiд впливом гiдролiзу глiкоалкалоiди розпадаються на цукор i аглiкон, фiзичнi i хiмiчнi властивостi якого i вказують на велику схожiсть його зi стероiдами. Стероiди утворюють рiзноманiтну групу важливих природних речовин. Сюди вiдносяться стерiни, жовчнi кислоти, гормони розмноження, вiтамiни групи Д, глiкозиди, рiзнi алкалоiди i iншi речовини. Базовою структурою усiх стероiдiв i стеран (циклопентанопергiдрофенантрен).
До класу стероiдiв належать також фiтостерини: сiто-, стiгма-, i кампестерiн, холестерiн, ергостерiн, жовчнi кислоти.
Сполуки стероiдiв з сахаром належать до групи глiкозiдiв. В цю групу, крiм глiкоалкалоiдiв, входять Влсердечнi глiкозiдиВ», якi мiстяться в деяких рослинах iз родини Strophanthus i Digitalis. Стероiднi саконiни також являють собою глiкозiди аглiконiв, котрi э похiдними циклопентанопергiдрофенантрена. Сапонiни не мiстять в своiму складi атом азоту [ ].
Одним з доказiв схожостi аглiкона глiкоалкалоiдiв зi стероiдами i те, що iх можливо перетворити в метилциклофенантрен. Ця сполука синтезуiться тим же шляхом iз стероiдiв. При третьому атомi вуглецю аглiкона знаходиться група, яка може сполучатися глiкозiдно з моносахаридами. Остаточно стероiдну будову аглiкона вдалося довести, коли похiдне аглiкона отримали з вiдомих стероiдiв i, навпаки, похiднi стероiдiв- iз аглiкона глiкоалкалоiдiв; наприклад, iз безазотноi речовини криптогенiна - соласодiн, iз соласодiна - стероiдний сапогенiн - дiосгенiн. Велiка схожiсть з сапонiнами виявляiться i в будовi сахарноi частини глiкоалкалоiдiв. Загальною властивiстю сапонинiв i глiкоалкалоiдiв являiться iх гемолiтична дiя [ ].
Таким чином, основними глiкоалкалоiдами культурноi картоплi i a- соланiн та a-чаконiн. Всi глiкоалкалоiди рiзних видiв картоплi складаються з двох основних частин: аглiкона (алкалоiда), який маi стероiдну будову, i молекули сахару, в склад якоi входять моно-, дi-, три-, або тетрасахариди.
1.1.2. Глiкоалкалоiди як токсичнi сполуки
Глiкоалкалоiди соланiн та чаконiн в невеликiй кiлькостi розглядаiться як цiлком можливi компоненти бульб картоплi, при надмiрнiй кiлькостi вони погiршують смаковi якостi картоплi. У бульб з`являiться гiркуватий смак, при вживаннi таких бульб в харчування вiникають чуття зуду в горлянцi [ ]. Встановлена визначена кореляцiя мiж наявнiстю глiкоалкалоiдiв i органолептичними смаковими якостями вареноi картоплi. При дуже низькому вмiстi глiкоалкалоiдiв в бульбах (порядку 1-5 мг%) смак вареноi картоплi трохи гiрший, нiж при середньому (5-10 мг%). Але збiльшення iх вмiсту вище за 26 мг% на сиру речовину дуже погiршуi смак вареноi картоплi i стаi небезпечним для здоров`я людини [ ].
Бульби картоплi, якi мiстять нiж 20 мг% глiкоалкалоiдiв, не рекомендуiться вживати в харчування, осбливо коли картопля вареться зi шкiрочкою. Почищенi бульби менш небезпечнi, так як значна частина глiкоалкалоiдiв видаляiться зi шкiрочкою, а частина переходить в воду, при вариннi ж неочищених бульб цього не вiдбуваiться вз`язку з слабкою проникнiстю шкiрочки. За даними М.Г.Шевченко [ ], нйбiльшу небезпеку для здоров`я становить нерозчинний в водi соланiн, зосереджений в основi пророслх вiчок.
1.1.3. Роль глiкоалкалоiдiв в явищi iмунiтета до бактерiальних, грибкових захворювань i шкiдникiв та iх значення в селекцii нових сортiв картоплi.
Глiкоалкалоiди привертають увагу селекцiонерiв-картоплярiв в зв`язку з iх токсичною дiiю на бактерiальнi та грибковi захворювання картоплi, а також в зв`язку з токсичною дiiю на колорадського жука [ ].
Встановлено, що соланiн затримуi рiст бактерiй, а 0,1-0,5% -ний розчин його викликаi вiдмирання протоплазми. Видiленi фунгiциднi якостi глiкоалкалоiдiв в вiдношеннi ряду патогенних грибiв, таких як Microsporum audoine, Trichophyton gypseum, Fusarium oxуsporum, зооспор Phytophthora infestans, Helminthosporum carbonum.
В РЖнститутi бiохiмii РАН дослiджували фунгiциднi властивостi глiкоалкалоiдiв i iнших сполук картоплi, надiлених антибiотичними якостями, а також вивчали дiю механiчних пошкоджень i зараження Phytophtora infestans на утворення i розподiл глiкоалкалоiдiв в тканинах бульб [ ]. Виявилось, що глiкоалкалоiдам a-соланiну та a-чаконiну притаманна бiльша фунгiцидна активнiсть нiж фенолам (кофейнiй i хлорогеновiй кислотам). Так, в концентрацii 0,1 мг/мл останнi не впливали на кiнетичнi показники росту фiтофтори, тодi як глiкоалкалоiди пригнiчували його майже на 70%. Зооспори цього паразита гинули вже при концентрацii a-соланiна 5 мкг/мл.
Соланин, чаконiн i соланiдiн iнгiбують радiальний рост Alternaria solani на картоплi. По ступеню iнгiбування на першому мiсцi стоiть соладiнiн, потiм чаконiн i соланiн. При концентрацii 500 мг/л соланiн викликаi 50% iнгiбування росту пiсля 96-годинного перiоду iнкубацii при 24оС. Встановлено наявнiсть збiльшення чутливостi листкiв до розвiтку пошкодження яке супроводжуiться зменшенням концентрацii глiкоалкалоiдiв в листках в мiру iх старiння. Наприклад, 30-деннi листки сорту Катадин мiстять 950 мг/кг глiкоалкалоiдiв i стiйкi до паразиту, тодi як старiючi 90-деннi листки мiстять тiльки 150мг/кг глiкоалкалоiдiв i значно уражаються грибом [ ].
Проте подальшi дослiдження показали, що хоча глiкоалкалоiди i надiленi фунгiтоксичнiстю, вони все ж таки не i вирiшальним фактором в явищi фiтоiмунiтета картоплi [ ]. Взагалi пiсля iнфiкування в бульбах картоплi накопичуiться хлогенова та кофейна кислоти, скополетин, a-соланiн, a-чаконiн, рiшитин, фiтуберин. Всi зазначенi сполуки, за виключенням рiшитiна та фiтуберiна, присутнi в шкiрочцi бульби в кiлькостях рiвних або бiльших, анiж синтезованих за умов iнфiкування патогеном очищеноi картоплi. Хлорогенова та кофейна кислоти знайденi в усiх частинах рослини. Вони продукуються в бульбах картоплi в результатi пошкоджень та iнфекцii багатьма патогенними та не патогенними органiзмами i накопичуються (або присутнi) в деяких здорових тканинах рослини картоплi в кiлькостях, якi можуть пригнiчувати мiкроорганiзми. Механiчне пошкодження чи iнфекцiя викликаi швидке окислення цих речовин, а продукти окислення токсичнi для багатьох мiкроорганiзмiв. На раннiх стадiях iнфекцii пiсля iнокуляцii бульб деякими расами фiтофтори накопичуються полiфеноли. В тканинах, iнокульованих несумiсними расами фiтофтори, або в зрiзах накопичуiться тiльки хлорогенова кислота.
Глiкоалкалоiди a-соланiн i a-чаконiн зосереджуються бiля пошкоджених дiлянок бульби [ ]. Про те, що глiкоалкалоiди не i вирiшальним фактором в iмунiтетi картоплi, свiдчать результати дослiдiв, проведених на раневiй перидермi бульб. В зараженiй фiтофторою раневiй тканинi картоплi глiкоалкалоiди не накопичуються. Звiдси виходить припущення, що фiтофтора надiлена здатнiстю руйнувати глiкоалкалоiди.
В раневiй тканинi, зараженiй несумiсною расою фiтофтори, не вiдбуваiться накопичення глiкоалкалоiдiв, не виявляються фiтоaлексини рiшитин i любiмiн. В непошкодженiй паренхимi бульб не синтезуються нi глiкоалкалоiди, анi фiтоалексiни. В раневiй перидермi весь мевалонат використовуiться на синтез глiкоалкалоiдiв. В вiдмерлiй тканинi, навпаки. Утворення глiкоалкалоiдiв майже не вiдбуваiться, а мевалонат використовуiться для синтезу рiшитiна та любiмiна. В зараженiй тканинi, як i в раневiй, глiкоалкалоiди хоча i утворюються, але розкладаються фiтофторою [ ].
В США проводили роботи по виведенню сортiв картоплi, надiлених полiгенною стiкiстю до фiтофтори, при цьому стикалися з труднощами, обумовленими отриманням гiбридiв з високим вмiстом в бульбах глiкоалкалоiдiв. Дослiдження, проведенi в селекцiйному центрi штата Захiдна Вiргiнiя, на 15 клонах картоплi, надiлених рiзною ступеню полiгенноi стiйкостi до фiтофтори i трьох стандартних сортах, не пiдтвердили наявнiсть взаiмозв`язку мiж зазначеною стiйкiстю i вмiстом в бульбах глiкоалкалоiдiв. В результатi прийшли до висновку, що можливе створення нових сортiв картоплi з полiгенною стiйкiстю до фiтофтори i звичайним, або зниженим вмiстом глiкоалкалоiдiв в бульбах. Встановлена висока ступiнь кореляцii (r=0,82) мiж вмiстом глiкоалкалоiдiв в листках та бульбах випробуваного селекцiйного матерiалу [ ].
Дослiдженнями Куна було встановлено, що непоiдання листкiв дикого виду картоплi Solanum demissum колорадським жуком обумовлена наявнiстю в них глiкоалкалоiда демiсiна [ ]. Це спонукало вiтчизняних ботанiкiв органiзувати експедицiю в краiни Пiвденноi Америки для пошукiв диких видiв картоплi, стiйких до колорадського жука. Пiд час експедицii було знайдено бiля 10 таких видiв, частина з них використовуiться в селекцii при виведеннi нових сортiв картоплi.
Глiкоалкалоiди бульбоносних видiв дикоi картоплi належать головним чином до групи соланiну чи демiсину. Найбiльша кiлькiсть стiйких до колорадського жука видiв картоплi тАУ S.commersonii, S.chacoense, S.gibberulosum, S.shikii, S.dolichostigma вiдноситься до Commersonia, але демiсин в них вiдсутнiй. Високостiйкими до колорадського жука виявились також S.polyadenum, S.jamesii, котрi мiстять малу кiлькiсть демiсину. Дикi види картоплi, якi характеризуються низьким вмiстом демiсину, за виключенням видiв Acaulia, виявились надто стiйкими до колорадського жука [ ].
2. ПРИРОДНРЖ УМОВИ ГОСПОДАРСТВА
Виробнича практика була проведена в учбово-навчальному комплексi (УНВК) Сумського державного аграрного унiверситету (СДАУ) який знаходится в м. Суми (пiвденна частина). УНВК розташований у пiвнiчнiй частинi Лiвобережного Украiнського Лiсостепу, яка в цiлому характеризуiться помiрним клiматом. Лiто тАУ тепле, iз значною кiлькiстю опадiв. Зима тАУ не дуже холодна, з вiдлигами.
Згiдно багаторiчних даних Сумськоi метеостанцii, середньорiчна температура району, де розташований УНВК, дорiвнюi 60 С (табл. 1). Найбiльш холодними мiсяцями i сiчень i лютий, найбiльш теплими тАУ липень, при чому абсолютний мiнiмум температур повiтря -360 С спостерiгаiться в сiчнi, а максимум +380 С - у серпнi. Останнi веснянi заморозки у повiтрi спостерiгаються у РЖРЖРЖ декадi квiтня, в окремi роки тАУ на початку травня, а першi осiннi заморозки тАУ на початку жовтня, з
1. Середня багаторiчна температура повiтря за мiсяцями
| Мiсяцi | РЖ | РЖРЖ | РЖРЖРЖ | IV | V | VI | VII | VIII | РЖХ | Х | ХРЖ | ХРЖРЖ | Середньо- рiчна |
| Середньо-мiсячна температура повiтря | -7,8 | -7,6 | -2,6 | 6,3 | 14,6 | 17,4 | 19,6 | 18,3 | 13,0 | 6,6 | -0,1 | -5,5 | 6,0 |



 коливаннями 11.09. тАУ 27.10. Довжина безморозного перiоду складаi приблизно 155 днiв. Зима снiжна, характеризуiться нестiйкою погодою, поряд iз низькими температурами -250 - -300 спостерiгаються вiдлиги з температурами +40 - +50.
коливаннями 11.09. тАУ 27.10. Довжина безморозного перiоду складаi приблизно 155 днiв. Зима снiжна, характеризуiться нестiйкою погодою, поряд iз низькими температурами -250 - -300 спостерiгаються вiдлиги з температурами +40 - +50.
Частi вiдлиги взимку при глибокому промерзаннi грунту iнодi призводить до застою талих вод i утворенню льодяноi кiрки, що негативно впливаi на перезимiвлю озимих. Висота снiгового покрову досягаi 22 см. Розподiлення його на бiльшiй частинi територii нерiвномiрне: снiг накопичуiться у балках, ложбинах стоку.
Максимальна глибина промерзання грунту на рiвних дiлянках сягаi 135 см, мiнiмально тАУ 40 см, середня тАУ 88 см.
Середньорiчна кiлькiсть опадiв становить 510 мм (табл. 2).
2. Середня багаторiчна кiлькiсть опадiв за мiсяцями
| Мiсяцi | РЖ | РЖРЖ | РЖРЖРЖ | IV | V | VI | VII | VIII | РЖХ | Х | ХРЖ | ХРЖРЖ | За рiк |
| Опади в мм | 28 | 23 | 26 | 40 | 43 | 67 | 68 | 53 | 45 | 42 | 33 | 37 | 510 |
З таблицi видно, що найбiльша кiлькiсть опадiв випадаi у лiтньо-весняний перiод, що спiвпадаi з максимальним ростом сiльськогосподарських культур i сiвбою озимих i сприяi iх розвитку.
Взимку переважають пiвнiчно-схiдний та пiвнiчно-захiдний вiтри. Напрямок iх часто змiнюiться, що призводить до рiзних змiн температури. За останнi роки спостерiгаються суховiйнi пiвденно-схiднi вiтри, якi сильно висушують грунт i знижують врожай основних сiльськогосподарських культур.
Рельiф УНВК рiвнинний, але захiдна частина дещо полога i засiваiться багаторiчними травами.
Грунти, якi представленi на дослiдному полi, -це типовi чорноземи, важкосуглинистi, середньогумуснi на льосовидному суглинку. Грунт характеризуiться високим вмiстом крупнопилови часток (0,01-0,05 мм), частка яких сягаi 55% i бiльше. Вмiст часток, якi належать до фракцii розмiром 0,001 мм i менше сягаi 25-30%, що обумовлюi високу iмнiсть поглинання, яка досягаi 28-30 мг/екв на 100 г грунту. Реакцiя грунту практично нейтральна. Значення рН коливаiться в межах 6,5-7,5/6,5-7,6 у верхнiх горизонтах i 7,3-7,5 у нижнiх (карбонатних).
Розорювана частина грунту приблизно до 30 см., яка мiстить у собi необхiднi елементи живлення (N, P, K) : азоту нiтратного 2,2-3,; азота амонiйного 11,2-10,6; фосфора 157, калiя 70 мг на 100 г грунту, вiдсоток гумусу 4,0%.
УНВК заснований в 1987 р. для проведення науково-дослiдноi роботи та для демонстрацii рiзних технологiй вирощування сiльськогосподарських культур. До складу УНВК входять комплекс по оздоровленню насiнневоi картоплi методами меристеми, обласне обтАЩiднання тАЬСортнасiняовочтАЭ i науково-дослiдне обтАЩiднання тАЬЕлiтатАЭ з лабораторiiю картоплi i дослiдним господарством. До складу господарства також входять дослiднi поля в с.Постольне, Гамалiiвка та Косiвщина, де вирощуються озима пшениця, овес, гречка, картопля, ячмiнь, овес, гречка, горох, багаторiчнi трави, капуста, морква, столовi буряки, цибуля. Урожайнiсть деяких культур в 2000 роцi: озима пшениця 35 ц/га; овес 41 ц/га; гречка 20 ц/га; картопля 150 ц/га; соняшник 15 ц/га; яра пшениця 13 ц/га; горох 13 ц/га; буряки цукровi 460 ц/га; кукурудза на зерно тАУ 42 ц/га.
Дослiднi поля УНВК займають 43 га сiльськогосподарських угiдь. Сiвозмiна займаi майже всi 43 га. Схема сiвозмiни така:
1. Овес 10 га.
2. Озима пшениця 5 га.
3. Картопля (насiння )0,7.
4. Ячмiнь .
5. Гречка 4,6 га.
6. Цукровий буряк 10 га.
РЖз сiвозмiни видiлена дiлянка 2,5 га, розмiщена на схилi, яка засiваiться багаторiчними травами для тварин вiварiя.
Крiм того в межах УНВК вирощували в 2000 р.такi культури:
1. Багаторiчнi трави 2,4 га
2. Молодий сад 3 га
3. Томати 0,2 га
4. Цибуля 0,25 га
5. Буряки столовi 0,07 га
6. Морква 0,07 га
7. Крiп 0,01 га
8. Петрушка 0,01 га
9. Капуста 0,75 га
10. Огiрки 0,05 га.
На дослiдних дiлянках проводять експериментальнi роботи кафедри селекцii (0,7 га) i рослинництва (0,3 га).
Для роботи на дослiдних полях УНВК закрiплений пiдроздiл i повний комплекс тракторiв та сiльськогосподарського обладнання. Для проведення дослiдiв видiлено дiлянки площею в 0,1 га. В склад пiдроздiлу входять: завiдуючий дослiдним полем i два iнженери. Для зберiгання i ремонту технiки за дослiдним полем закрiплений спецiальний бокс, а для зберiгання зерновоi продукцii - ангар. Технiка УНВК представлена тракторами Т-150, Т-70, Т-150К, МТЗ-82, Т-25, Т-16.
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)