Влияние цитокинина на рост и развитие проростков пшеницы в зависимости от условий минерального питания
МИНЕСТЕРСТВО ОБРАЗОВАНИЯ РФ
МОСКОВСКИЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ
Естественно тАУ экологический фтАУт
Кафедра биологии и экологии организмов
Стахович Артем Валентинович
Д И П Л О М Н А Я Р А Б О Т А
Тема: тАЬВЛИЯНИЕ ЦИТОКИНИНОВ НА РОСТ И РАЗВИТИЕ
ПРОРОСТКОВ ПШЕНИЦЫ В ЗАВИСИМОСТИ ОТ
УСЛОВИЙ МИНЕРАЛЬНОГО ПИТАНИЯтАЭ.
Научный руководитель: Кандидат биологических наук, доцент
ТАРАСЕНКО АЛЛА АНДРЕЕВНА
Москва 2000 год
СОДЕРЖАНИЕ
| | Стр. |
| 1. | ВВЕДЕНИЕ | 3 |
| 2. | ОБЗОР ЛИТЕРАТУРЫ | 4 |
| 2.1 | Краткая характеристика фитогормонов | 4 |
| 2.2 | Общая характеристика цитокининов | 6 |
| а) | Химическая структура | 6 |
| б) | История открытия | 7 |
| 2.3 | Физиологическая роль цитокининов | 9 |
| а) | Стимуляция деления клеток | 9 |
| б) | Влияние цитокининов на рост клеток | 11 |
| в) | Действие цитокининов на органогенез | 13 |
| г) | Снятие с помощью цитокининов апикальной доминанты | 15 |
| д) | Прерывание покоя и стимуляция прорастания семян под действием цитокининов | 16 |
| е) | Влияние цитокининов на рост целых растений | 17 |
| ж) | Продлениес помощью цитокининов жизни срезанных листьев | 18 |
| з) | Цитокинины как фактор воздействия одних органов растения на другие и их влияние на передвижение веществ в растении | 18 |
| и) | Защитное действие цитокининов против неблагоприятных факторов | 19 |
| 2.4 | Роль калия в минеральном питании растений | 21 |
| а) | Источники калия для растений | 21 |
| б) | Значение калия в жизни растения | 23 |
| в) | Физиологическая роль калия в растениях | 23 |
| 3. | ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ | 27 |
| 3.1 | Характеристика объекта исследования | 27 |
| а) | Классификация | 27 |
| б) | Краткая ботаническая характеристика | 27 |
| в) | Характеристика сорта Терция | 28 |
| г) | Агрономическое значение яровой пшеницы | 29 |
| 3.2 | Методика исследования | 29 |
| 4. | РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ | 30 |
| 4.1 | Влияние калия на рост и физиологические процессы | 30 |
| 4.2 | Влияние кинетина на физиологические процессы | 34 |
| 4.3 | Выводы | 47 |
| 5. | СПИСОК ЛИТЕРАТУРЫ | 48 |
1. ВВЕДЕНИЕ
В последнее время при возделывании различных сельскохозяйственных культур все большее внимание уделяется приемам, с помощью которых можно воздействовать непосредственно на растительный организм. К таким приемам относится обработка растений или их семян различными веществами, в частности регуляторами роста.
Не смотря на то, что растение обладает способностью синтезировать гормоны, во многих случаях добавление их извне оказывает на растение положительное действие. Влияние растительных гормонов или их синтетических заменителей проявляется особенно резко тогда, когда уровень содержания их в растении не высок.
В настоящее время общепризнанно, рост и развитие растений регулируется системой фитогормонов, включающей ауксины, гибберелины, кинины и ингибиторы. Каждая группа гормонов имеет свои специфические функции. Фитогормоны по-разному реагируют на различные питательные смеси.
Задачей настоящей работы было исследование влияния цитокининов на рост и некоторые физиологические процессы растений пшеницы на фоне полной питательной смеси, при недостатке калия, и с внесением цитокинина.
2. ОБЗОР ЛИТЕРАТУРЫ
2.1 Краткая характеристика фитогормонов
Важнейшими представителями эндогенных регуляторов роста растений являются фитогормоны. Это - нормальные продукты жизнедеятельности самого растения, участвующие в регуляции обмена веществ и формообразовательных процессов на всех этапах онтогенеза (Муромцев Г.С., Агнистикова В.Н, 1984 ). Они обладают тремя общими основными свойствами:
1. Гормоны синтезируются в одном из органов растения (молодые листья, почки, верхушки корней и побегов) и транспортируются в другие места, где активируют процессы онтогенеза и роста.
2. Гормоны синтезируются и функционируют в растениях в микро количествах.
3. Гормоны могут вызывать характерные морфологические изменения.
Следовательно, этим веществам можно дать другое, более обобщенное, определение. Фитогормоны - это вещества, действующие в ничтожных количествах, образующиеся в одних органах и оказывающие регулярное влияние на какие-либо физиологические процессы в других органах растения.
Работы многих других ученых ( Чайлахян, Саркисова, Полевой, 1982 ) показали, что система гормональной регуляции во многом определяет характер протекания таких важнейших физиологических процессов, как рост, формирование новых органов, переход растений к цветению и формированию пола цветков, старение листьев, переход в состояние покоя и выход из него почек, клубней, луковиц и т.д.
Процессы роста и морфогенеза тоже являются физиологическими программами, только медленно протекающими. Учитывая выше сказанное, можно дать еще одно определение фитогормонам. Фитогормоны - соединения, с помощью которых осуществляется взаимодействие клеток, тканей и органов и которые необходимы в малых количествах для запуска и регуляции физиологических и морфогенетических программ.
В настоящее время известно четыре группы фитогормонов:
ауксины, гибберелины, цитокинины и ингибиторы роста. Все они могут активизировать, но могут и тормозить функциональную активность клеток. Общим условием для действия любого фитогормона является наличие в клетках специфичных рецепторов. Все фитогормоны вызывают у компетентных клеток сравнительно быстрые физиологические реакции, связанные, очевидно, с мембранами и более медленные изменения, зависящие от синтеза белков и нуклеиновых кислот ( Полевой В., 1982 ).
Помимо естественных фитогормонов, получено большое количество синтетических аналогов этих природных соединений, которые часто обладают высокой физиологической активностью. Строго говоря, эти вещества не могут отнесены к фитогормонам, так как не образуются в растениях, однако многие из них по активности не уступают фитогормонам или даже превосходят их ( Муромцев Г.С., 1973 ).
2.2 Общая характетистика цитокининов
а) Химическая структура
Цитокининами называется один из типов фитогормонов, обладающих определенной совокупностью биологической активности, которая весьма разнообразна и проявляется при регуляции роста, органообразования, процессов старения и покоя.
Цитокинины принимают участие наряду с другими фитогормонами в регуляции самых разнообразных физиологических процессов в растении. Для них, как и для других фитогормонов, характерна полифункциональность. Кроме того, действие цитокининов появляется в неразрывной связи с действием других фитогормонов и природных ингибиторов.
В химическом отношении природные цитокинины и их синтетические заменители представляют собой производные
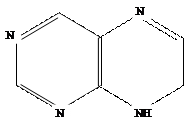 6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
Пурин
Цитокинины пуринового ряда слаборастворимы в воде, но хорошо растворимы в этаноле, этиловом эфире, ряде щелочей и кислот. Цитокинины устойчивы к нагреванию, автоклавированию, действию щелочей и кислот.
Однако точная химическая характеристика цитокининов, как производных 6-аминопурина, затрудняется тем, что такими же свойствами обладает также дифенилмочевина и ее многочисленные производные. Очевидно, вопрос о химической характеристике цитокининов получит окончательное разъяснение только после установления молекулярного механизма их действия.
б) История открытия
Цитокинины были открыты в 1955 г. Скугом и Миллером с сотрудниками в Висконсинском университете в США. Их обнаружению помог так называемый тАЬдефектный объекттАЭ, которым явился каллюс сердцевины стебля табака. Он образовался на кусочках сердцевины стебля в условиях стерильной культуры, но быстро прекращал рост в связи с исчерпанием какого-то фактора, исходно присутствовавшего в тканях стебля. Ввиду того, что вернуть активный рост каллюса удавалось добавлением к питательной смеси, содержащей индолилуксусную кислоту ( ИУК ), дрожжевого экстракта, а так же кокосового молока, Скугом и сотрудниками была предпринята попытка выделить недостающие для роста каллюса вещество из дрожжевого экстракта. При этом выяснилось, что активное вещество обладает свойствами пурина.
Вместе с тем проверка природных пуриновых оснований так же как гидрометода РНК и ДНК показала, что они не способны обеспечить рост стеблевого каллюса табака. Неожиданно активным оказался эфирный экстракт из старого, давно хранившегося препарата ДНК. Впоследствии выяснилось, что активное вещество образуется в любом препарате ДНК при ее деградации путем автоклавирования в кислой среде. Это вещество было выделено в виде кристаллов и идентифицировано химически. Оно оказалось 6- фурфуриламинопурином.
6-фурфуриламинопурин
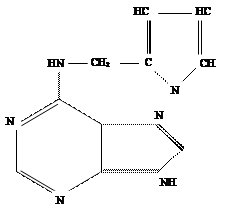
Указанное соединение образуется при автоклавировании ДНК в кислой среде из дезоксиаденозина. Поскольку 6-фурфуриламино-пурин в присутствии ИУК обеспечивал клеточное деление в изолированной сердцевине ткани стебля табака, он был назван кинетином ( от слова тАЬкинезтАЭ ).
Принцип химического синтеза производных пурина с замещением в аминогруппе у шестого атома углерода пуринового кольца был известен ранее, поэтому вслед за химической идентификацией кинетина последовал его синтез. Затем было синтезировано много других активных соединений, которые отличались от кинетина характером заместителя в аминогруппе при шестом атоме углерода. Все эти соединения, включая кинетин, были объединены под общим названием кинины. Это название было распространено в литературе до 1965 г. Однако ввиду того, что в физиологии животных термин тАЬкининытАЭ еще ранее был применен для совершенно иных соединений - биологически активных
полипептидов, Скуг, Стронг и Миллер предложили заменить название кинины на цитокинины. В настоящее время термин цитокинины стал общепризнанным.
2.3 Физиологическая роль цитокининов
а) Стимуляция деления клеток
Цитокинины были открыты как вещества, необходимые для деления клеток у стеблевого каллюса табака и изолированной сердцевинной ткани стебля табака (Мi11еr, 1956; Skoog, Мil1еr, 1957). Эти ткани не способны к синтезу цитокининов, а также ауксинов, что позволило выявить многие существенные стороны действия данных фитогормонов на ростовые процессы. Так, в работах Скуга с сотрудниками было показано, что у изолированной сердцевинной ткани стебля табака в стерильной культуре одна ИУК несколько усиливала синтез ДНК, вызывала в отдельных клетках митозы, но не индуцировала клеточных делений. Точно так же один кинетин не вызывал деления клеток. В его присутствии не происходили митозы, а синтез ДНК он стимулировал в меньшей степени, чем ИУК. Только совместное действие ауксинов и кинетина значительно активировало синтез ДНК, вызывало митозы и индуцировало деление, клеток в изолированной сердцевине стебля табака (Dаs еt а1., 1956, 1958).
В последующем были предприняты попытки разграничить во времени действие ИУК и кинетина в индукции клеточных делений у этой ткани (Nitsch, 1968; Дмитриева, 1972). Полученные данные не имеют пока однозначной интерпретации, но позволяют предполагать, что начальные стадии процесса индуцируются одним ауксином, тогда как в последующем необходимы оба гормона ( Дмитриева, 1972 ).
Таким образом, было установлено, что регуляция деления клеток в изолированной сердцевинной ткани стебля табака осуществляется не одним гормоном - гипотетическим индуктором клеточных делений, а одновременным действием двух гормонов - цитокинина и ауксина. При этом для каждого фитогормона был показан свой предел концентраций, в которых он при наличии в среде другого гормона стимулирует данный процесс. Например, кинетин в присутствии в питательной среде 2 мг/л ИУК стимулировал деление клеток сердцевинной ткани стебля табака, начиная с концентрации 0,005 мг/л, номаксимальное его действие достигалось в пределах концентраций от 0,02 до 0,5 мг/л. Увеличение концентрации кинетинадо 2 мг/л приводило к угнетению процесса. Стимулирующее действие ИУК при одном и том же содержании в среде кинетина нарастало с увеличением ее концентрации до 1,8 мг/л. Максимальную интенсивность деления клеток вызывало сочетание оптимальных концентраций в среде обоих фитогормонов ( Scoog , Miller, 1957 ) .
Вместе с тем в литературе появилось сообщение о том, что если заменить ИУК на2,4-дихлорфеноксиуксусную кислоту (2,4-Д) и взять последнюю в очень высокой концентрации, можно получить слабо идущее деление клеток сердцевинного каллюса табака и в отсутствие кинетина ( Witham,1968). По поводу этих данных возникают два предположения: либо только ауксины критичны для деления клеток каллюса, либо высокие концентрации 2,4-Д индуцируют синтез в каллюсе эндогенных цитокининов. Дальнейшие исследования должны показать, какое из этих предположений справедливо.
В связи с этим важно упомянуть, что у отдельных штаммов каллюса сердцевины табака обнаружена способность к синтезу цитокининов. Эти штаммы возникали спонтанно при длительной культуре каллюса (Fох 1963). Одни из них были способны к синтезу только цитокининов, а другие тАУ и цитокининов и ауксинов. Для интенсивного роста таких штаммов не требовалось добавления в питательную среду соответствующих фитогормонов ( Fox, 1961 ). Они сами могли служить источником фитогормонов для обычных дефектных штаммов. Аналогичное явление наблюдалось и для каллюсов семядолей сои, которые обычно также не способны к синтезу ауксинов и цитокининов и нуждаются для роста в их предоставлении извне (Miller, 1963). В месте с тем были обнаружены штаммы этой ткани, не зависящие в своем росте от присутствия в среде цитокининов, и было показано, что это происходит потому, что они приобретают способность к синтезу собственных цитокининов ( Miura, Miller, 1969 ).
Достаточное эндогенное содержание цитокининов, нехватка ауксинов или других необходимых веществ могут стать причиной отсутствия стимулирующего действия цитокининов на деление клеток. Кроме того, в случае высокого эндогенного содержания цитокининов их предоставление извне может привести и к угнетению клеточных делений. Возможно именно с таких позиций следует рассматривать работы, в которых показано отсутствие стимуляции или даже торможение деления клеток цитокининами ( Humphries, Wheeler, 1960; Mc Manus, 1960; Vant Hof, 1968). Кроме того нужно еще иметь в виду, что некоторые растительные объекты могут быть не компетентны для реакции на цитокинин активацией клеточных делений.
Вместе с тем стимуляция с помощью цитокининов митозов и деления клеток продемонстрирована на очень большом числе разнообразных объектов, включая изолированные ткани (Мi11ег, 1961а; Syono, 1965; Neumann еt а1., 1969; Fosket, Тоrrеу, 1969), кончики корня ( Guttman, 1956; Olszewska, 1959а, b), растущие листья (Саtarino, 1965), точки роста стебля (Саtarino, 1965), зародыши прорастающих семян ( Haber, Luippold 1960а, b), водоросли (Мoewus, 1959), мхи ( Szweykowska еt а1., 1968) и другие объекты ( Guttman, 1957; Olszewska, 1959b; Walker, Dietrich, 1961; Мi11еr, 1961а).
б) Влияние цитокининов на рост клеток
Уже в первых работах по цитокининам на стеблевых каллюсах табака в лаборатории Скуга было показано, что цитокинины влияют не только на деление клеток, но и на их рост растяжением. Скуг пришел к выводу о том, что для роста клеток так же, как и для их деления, нужны и цитокинины, и ауксины, причем концентрации этих веществ и соотношение между ними, стимулирующее рост клеток, отличается от тех, которые стимулируют клеточное деление ( Scoog, Мil1еr, 1957).
Наиболее эффективно цитокинины стимулируют рост клеток в отрезках растущих листьев. Это было показано на листьях самых. различных двудольных травянистых растений ( Scott, Liverman, 1956; Мi11еr, 1956; Powell, Griffith , 1960, 1963; Кuraishi, Okumura, 1956; Кuraishi, 1959; Сatarino, 1965), а также на семядолях (Ваnerji, Laloraya, 1965, 1966; Курсанов и др., 1969).
На отрезках листьев однодольных растений стимуляция роста клеток цитокинином не удается (Кuraishi, 1959). С помощью цитокинииов удается индуцировать рост клеток в листьях, уже давно закончивших рост (Кuraishi, 1959; Моthes, 1960). Помимо листьев, стимуляция роста клеток цитокининами показана и у многих других объектов ( Schrank, 1958; Саtarino, 1965;. Аrora еt а1., 1959; Wardlaw, Мitга, 1958; Мi11еr, 1961а).
Однако в литературе отмечены случаи, когда цитокинины угнетали рост клеток. Угнетение чаще всего обнаруживается на отрезках стебля и корнях (Мi11еr, 1961а; dе Rорр, 1956; Deysson, 1959; Butcter, Street, 1960; Вanerji, Lа1оrауа, 1967; Sprent, 1968b; Gaspar еt а1., 1969), причем цитокинины снимают стимулирующее действие на рост стебля ауксинов ( Brian, Hemming, 1957; Vanderhoff, Кеу, 1968) .
Однако не исключено, что различие роста клеток листьев и клеток стебля и корня при воздействии цитокининов определяется различием в диапазоне стимулирующих концентраций. Такое предположение основывается на работах, в которых использование очень низких концентраций цитокининов приводило к стимуляции роста стебля ( Supniewski еt а1., 1957; Sуоnо, Yamaki, 1963b). Этот вопрос подлежит специальному изучению.
в) Действие цитокининов на органогенез
Индукция с помощью цитокининов органогенеза у недифференцированной ткани стеблевого каллюса табака была впервые показана Скугом с сотрудниками, которые с помощью ИУК и кинетина вызывали образование у каллюса корней и побегов и установили, что для закладки каждого из, этих органов требовались свои специфические концентрации обоих фитогормонов. Меняя содержание фитогормонов в питательной среде, можно было направлять органогенез в сторону образования корней или побегов ( Scoog, Мi11еr, 1957).
Так, например, содержание в питательной среде 2 мг/л ИУК и 0,02 мг/л кинетина вызывало у каллюса сердцевинной ткани стебля табака интенсивное деление клеток и дифференциацию корней. Повышение концентрации кинетина до 0,5 тАУ 1 мг/л угнетало корнеобразование, но индуцировало формирование стеблевых почек. Дальнейшее увеличение концентрации кинетина до 2 мг/л оказывало общее ингибирующее действие на рост каллюса и процессы органообразования ( Scoog, Мi11еr, 1957).
Стимуляция с помощью цитокининов процессов органогенеза была затем подтверждена на очень многих объектах ( Gorton et al., 1957; Тоrrеу, 1958; Рrovasoli, 1958; Schrandolf, Reinert, 1959, Мitra, Allsopp , 1959; Бутенко, Яковлева, 1962; Мitra еt а1., 1962; Бутенко, 1964; Неide, 1965; Саtarino, 1965; Бутенко, Володарский, 1967; Вrandes, 1967; Ворр, Dickmann, 1967; Brandes, Кеndе, 1968; Реterson, 1969) .
При этом в ряде случаев цитокинины, стимулируя образование побегов, вместе с тем задерживали закладку корней. Однако Хайде на листьях бегонии показал, что закладка и корней, и стеблевых почек нуждается у этого объекта в цитокининах и ауксинах, и для каждого из этих процессов существуют оптимальные концентрации обоих фитогормонов, при которых они усиливают действие друг друга (Неidе, 1965а, b). Так как концентрации цитокининов, стимулирующие закладку корней, крайне низки ( Scoog,Мil1еr, 1957; Allsopp, Szweykowska, 1960), в большинстве объектов хватает, по-видимому, собственных цитокининов для осуществления этого процесса. Возможно, именно этим объясняется тот факт, что данные извне цитокинины ингибируют корнеобразование (Rорр, 1956; Sorokin еt а1., 1962; Nieuwhof, 1967). Вместе с тем у каллюса Рорulus tremuloides L. корнеобразование происходило только при полном удалении из питательной среды кинетина и при наличии в среде 2,4-Д. Кроме того, образование побегов требовало у этого объекта удаления из среды ауксинов и происходило под действием одного цитокинина ( Wolter, 1968). Эти данные противоречат результатам Скуга о необходимости обоих фитогормонов для дифференциации, как корней, так и побегов. Однако это противоречие могло бы быть разрешено с позиции большего эндогенного содержания обоих фитогормонов в каллюсе Рорulus tremuloides L., по сравнению с их содержанием в каллюсе сердцевины стебля табака. Этот вопрос требует дальнейшего изучения.
Существенно отметить, что с помощью цитокининов удается также влиять на закладку и дифференциацию генеративных органов, вызывая зацветание растений в условиях неблагоприятного температурного или фотопериодического режима (Чайлахян и др.,1961; " Negi, Olmo,1966; Маheshwari еt а1., 1966; Gupta, Маheshwari, 1969) . С помощью цитокинина можно также повлиять на пол цветков ( Negi, Оlmo, 1966) .
Цитокинины индуцируют органообразовательные процессы также у мхов (Skinner, Shive, 1955). Под действием цитокининов у протонемы мха, представляющей собою нитевидное образование, индуцируется закладка стеблевых почек ( Szweykowska, 1962, 1963, 1966; Szweykowska et al., 1968а, b, 1969, 1970; Szweykowska, Mackowiak, 1962; Szweykowska, Handszu, 1965; Szweykowska,Schneider,1967; Brandes, Kende,1968).
На основе индукцииобразования стеблевых почек у протонемы мхов разработан высокочувствительный биотест на цитокинины ( Hahn, Bорр, 1968). Известно также влияние цитокининов на процессы дифференциации у папоротников ( Gaudet, Нuang, 1967) .
Существенно, что действие цитокинина на процессы дифференциации обнаруживается у одноклеточной водоросли Аcttabularia ( Zetsche, 1963) . Эта гигантская клетка образует в ходе своего развития подобие корешка, стебля и шляпки. В зависимости от взятой концентрации кинетин стимулирует или угнетает образование и рост этих частей клетки. При этом стимулирующие концентрации для стебля и шляпки оказываются различными и меняются в зависимости от возраста клеток.
Стимулируя процессы дифференциации, цитокинины в ряде случаев вызывают де дифференциацию (МасNuff, von Maltzahn, 1960; Wardlaw, Mitra, 1958).
Таким образом, цитокинины оказывают большое влияние на процесс дифференциации, характер которого меняется в зависимости от концентрации цитокинина, его соотношения с ауксином и особенностей объекта.
г) Снятие с помощью цитокининов апикальной доминанты
По предположению Тиманна, цитокинины и ауксины участвуют в регуляции распределения ростовых процессов вдоль по стеблю растения. При
этом, если с помощью ауксинов осуществляется подавление роста боковых почек со стороны верхней почки и рост стебля происходит без образования боковых побегов (так называемая апикальная доминанта), то цитокинины снимают ингибирующее действие на латеральные почки верхушечной почки или ауксинов и вызывают рост боковых побегов ( Wickson, Тhimann, 1958, 1960; Lona, Восhi, 1957; von Maltzahn, 1959; Sebanek, 1965; Sachs, Тhimann, 1967; Sprent, 1968b). На этом основании можно с помощью цитокининов заменить главный побег на боковой ( Engelbrecht, 1967) или вызвать усиленный рост многих боковых побегов у растений, которые без обработки их раствором цитокинина не дают ветвления ( Lona, Восhi. 1957).
д) Прерывание покоя и стимуляция прорастания семян под действием цитокининов.
С помощью цитокининов удается прервать покой спящих почек древесных растений в летний и зимний периоды ( Сhvojka еt а1., 1961; Сhvojka еt а1., 1962; Вenes еt а1., 1965; Leike, 1967), а также вывести из состояния покоя клубни некоторых растений, например бегонии ( Esashi, Leopold, 1967).
Цитокинины оказываются эффективным средством в прерывании покоя некоторых семян ( Worsham е1 а1., 1959; Кhan, 1966). Стимулирующее действие цитокининов на прорастание обнаружено на семенах латука, которые нуждаются для индукции этого процесса в действии красного света (Мi11еr, 1956, 1958; Skinner et al., 1957; Haber, Luippold, 1960а, b; Ikuma, Thimann, 1963; Leff, 1964). Цитокинины стимулируют их прорастание как в темноте (Мi11ег, 1956), так и при кратковременном воздействии красным светом (Мil1еr, 1958) .
С помощью цитокининовудается повысить у ряда растений всхожесть семян, ослабленную в результате их длительного хранения (Курсанов и др., 1966а, б).
е) Влияние цитокининов на рост целых растений
Выше былиприведены примеры, когда с помощью цитокининов можно было вызвать рост боковых побегов на растении и их зацветание в неблагоприятных для цветения условиях. Что же касается общей стимуляции роста растений цитокининами, то ее получить весьма сложно и пока еще не найдены пути эффективного воздействия на рост и урожай растений. В этом отношении было сделано несколько неудачных попыток. Так, например, опрыскивание растений карликовой пшеницы раствором кинетина не влияло на их рост (Уао, Canvin, 1969), а полив растений томатов, росших в песке, раствором кинетина (10 тАУ 100 мг/л) резко угнетал их рост ( Kemp еt а1., 1957) . Кинетин, добавленный в питательную среду (0,002 тАУ 2 мг/л), тормозил также рост томатов, огурцов и гороха и задерживал их зацветание ( Witter, Dedolf, 1963). Неудачей закончились и наши попытки повлиять с помощью кинетина на формирование зерна и содержание в нем азотистых веществ у пшеницы (Курсанов и др., 1966а, б). По-видимому, во всех этих случаях в растениях хватало эндогенных цитокининов или, по крайней мере, не они являлись лимитирующим фактором.
Однако в литературе известны примеры и стимулирующего действия цитокининов на рост растений. Например, кинетин, добавленный в крайне низких концентрациях к раствору Кнопа, стимулировал рост проростков подсолнечника, фасоли и люпина ( Supniewski еt а1., 1957), растений ряски в темноте ( Hillman, 1957), проростков люпина в темноте и на свету ( Fries, 1960), а также проростков редиса на свету (Кuraishi, 1959). Погружение растений табака на 1 час корнями в раствор цитокинина (5*10-4 М) вызывало значительное усиление роста листьев на растении и повышало в них содержание калия (Dyar, 1968). Есть указания о возможности повлиять с помощью цитокининов нарост плодов яблони ( Letham,1969) и на формирование урожая у растений кукурузы (Власюк и др., 1969). Особенно интересные сведения получены в последнее время Турковой и Сабо (1970), которые показали, что однократное опрыскивание растений подсолнечника раствором кинетина на фоне высокой дозы азотного питания вызывает значительную прибавку общего веса растений и увеличение веса корзинок. Подобное же опрыскивание ускоряет и зацветание растений подсолнечника (Туркова, Фролова, 1970).
Приведенные примеры позволяют заключить, что, хотя случаи, когда эндогенные цитокинины бывают в растениях в минимуме и поэтому проявляется стимулирующее действие экзогенных цитокининов, бывают, по-видимому, редко, все жемогут быть найдены условия, при которых можно ожидать, что цитокинины окажутся эффективным средством воздействия на рост и урожай растений.
ж) Продление с помощью цитокининов жизни срезанных листьев
В предыдущих разделах показано участие цитокининов в регуляции роста и органообразования у растений. Однако, помимо этого цитокинины оказывают влияние на уже закончившие свой рост органы растения. Наиболее стильно их действие сказывается на срезанных листьях травянистых растений. Цитокинины задерживают их пожелтение и продлевают жизнь срезанных листьев.
з) Цитокинины как фактор воздействия одних органов растения на другие и их влияние на передвижение веществ в растении
Цитокинины, как и другие фитогормоны, являются одним из факторов, с помощью которого одни органы растения влияют на другие. Они оказывают регуляторное действие на распределение питательных веществ в растении.
и) Защитное действие цитокининов против неблагоприятных факторов
Цитокинины повышают устойчивость клеток к самым различным неблагоприятным воздействиям, таким как, действие повышенной и пониженной температуры, обезвоживание, гребная и вирусная инфекция, механическое воздействие и влияние различных химических агентов.
Так, в опытах Мотеса и Энгельбрехт было показано, что кинетин повышает устойчивость к кратковременному нагреву клеток обработанной им половины срезанного листа табака и способствует последующему репарационному процессу ( Моthes, 1960; Engelbrecht, Mothes, 1960, 1964 ). Это действие цитокининов Мотес ставит в прямую связь с их способностью повышать удерживание клетками подвижных метаболитов и приток к ним питательных веществ из окружающих тканей. Это свойство клеток удерживать и притягивать подвижные метаболиты Мотес называет аттрагирую щей способностью клеток. По данным Мотеса и Энгельбрехт, эта способность клеток листа резко снижается в результате нагрева, а цитокинин повышает ее, что может быть одним из путей проявления его защитного действия по отношению к нагреву. Механизм такого действия цитокининов не изучен.
Не исключено, что он связан с их влиянием на функциональное состояние мембран. Этот вопрос нуждается в специальном исследовании.
В связи со сказанным важно упомянуть, что защитное влияние цитокининов проявляется при действии на растительные объектысамых различных агентов, таких как, например, пенициллин ( Platt, 1958), хлорамфеникол (Моthes, 1960; Кулаева, Воробьева, 1962; Еngelbrecht, Nogai, 1964; Wollgiehn, Parthier, 1964), NaF и тетрахлорнитробензол ( Penot, 1963), 5-фторурацил ( Kim Won-Kyum, Greulach, 1961), 8-азагуанин (Мoewus, 1959; Вlaydes, 1966), токсические концентрации сахарозы ( Butcher, Street, 1960), высокий процент обогащения воды D2О (Соре еt а1., 1965), механическоевоздействие на клетки листа при инфильтрации ( Bagi, Farkas, 1968) и т. д. Причины защитного действия цитокининов в каждом конкретном случае могут быть различными и, очевидно, требуют специального изучения. Однако не исключено, что механизм такого действия может оказаться и одинаковым. Например, он может проявляться через действие цитокининов на структурное и функциональное состояние различных макромолекуляриых компонентов клетки и, в частности, на состояние их мембранного аппарата. Электронно-микроскопические исследования показывают, что цитокинины задерживают деградацию различных мембранных структур в клетках листа, включая ламеллы стромы и граны хлоропластов (Курсанов и др., 1964; Свешникова и др., 1966), мембранные структуры митохондрий (Курсанов и др., 1964) и эндоплазматический ретикулум (Shaw, Маnocha, 1965). При помощи цитокинина удавалось предотвратить разрушение в срезанных листьях мембранной оболочки сферозом, содержащих гидролитические ферменты, и таким путем защитить от разрушения белки, нуклеиновые кислоты и липоиды .плазмы (Ва1z, 1966). По-видимому, действие цитокининов на мембранный аппарат клетки может иметь существенное значение в осуществлении их регуляторного действия на обмен веществ растений и не исключено, что имеет отношение к защитному действию цитокининов при различных неблагоприятных воздействиях.
Иную интерпретацию может иметь защитное действие цитокининов на листья целых растений в условиях повышенной температуры (Mothes, 1960) и засухи (Моthes, 1961а; Shah, Loomis, 1965). По-видимому, в таких случаях в листьях может возникать дефицит эндогенных цитокининов за счет снижения их поступления из корней, что и создает условия для защитного действия данных извне цитокининов. Правда, необходимо упомянуть, что такое защитное действие проявляется не всегда. Так, при помощи цитокинина не удалось задержать пожелтение растений пшеницы при недостатке водоснабжения в условиях вегетационного опыта ( Nunes, 1967 тАУ 1968). Однако работа в этом направлении может быть перспективной для защиты растений от различных неблагоприятных воздействий.
2.4 Роль калия в минеральном питании растений
а) источники калия для растений
Калий представлен в природе тремя изотопами, из которых К39 составляет 93,31% , К40 - 0,01% , К41 - 6,68%. В свободном состоянии он не встречается в связи с большой химической активностью. Калий является сильным восстановителем. Он легко отдает свой внешний электрон и переходит при этом в одновалентный катион.
В почве находится в форме водно-растворимых солей, обменного и необменного калия, силикатов и алюмосиликатов. До 98% калия, находящегося в почве, недоступно растениям. Наиболее доступный и важный для растения - обменный калий. Растения поглощают калий из почвы.
Внутри растительной клетки калий распределен неравномерно.
Его не обнаружили в ядре и хлоропластах. Особенно много калия у молодых растений, т.е. в период, когда у растений идет интенсивное деление клеток и синтезируются органические вещества. Максимум калия у большинства растений отмечен к моменту цветения;особенно это характерно для злаков.
Физиологическая роль калия еще не достаточно ясна. Однако имеющийся экспериментальный материал позволяет сделать некоторые заключения о роли калия в растительных организмах. Важное значение имеет более или менее устойчивое соотношение концентраций ионов калия, натрия и кальция в организме. Калий, легко проникая внутрь клетки, увеличивает проницаемость клеточных мембран для различных веществ, чем оказывает значительное влияние на обмен веществ в самых разнообразных направлениях.
Процесс усвоения калия растениями из водных растворов зависит от состава анионов и катионов, входящих в питательную смесь в последнее время получены доказательства того, что анионы по степени своего влияния на интенсивность поглощения калия располагаются в следующий ряд:
HCO3 > CI > H2PO4 > NO3 > SO4
Катионы вступают в антагонистические взаимоотношения с калием, что сказывается определенным образом на процессе его поглощения. По степени проявления антагонизма катионы составляют следующий ряд:
Ca2+ > NH4+ > Na+ > Mg2+
Изучение взаимоотношений калия с другими ионами продолжается в связи с наличием многих противоречий по этому вопросу.
Процесс усвоения калия зависит от рН среды. Наиболее благоприятными являются слабо кислая или нейтральная реакция. При сдвиге рН в сильно кислую или щелочную сторону поглощение калия сильно тормозится ( Удовенко, Иванов, Ложкина, Урбанович, 1964 ).
В естественных условиях растения используют водно-растворимый калий, содержание которого в почвенном растворе пополняется за счет резервов почвенного поглощающего комплекса. Такой процесс идет на протяжении всей жизни растения ( Чириков, 1951 ).
Взаимосвязь растения с источниками калия в почве можно выразить схемой, приведены ниже.
Растение
^
калий почвы воднорастворимый
^
калий почвенного поглощающего комплекса,
выделенный в раствор
( Сa2+ , Mg2+ , Na+ , H+ )
б) значение калия в жизни растения
При недостатке калия в питательной среде у растений наблюдаются признаки калийного голодания. Наблюдается краевой запал листьев. У некоторых растений развивается хлороз, а у картофеля на листьях появляется бронзовый оттенок. Признаки голодания раньше проявляются у старых листьев, а потом и у более молодых. По-видимому, молодые листья могут некоторое время использовать запасы калия, накопленные в старых листьях, что свидетельствует о возможности повторного использования этого элемента. Семена, образовавшиеся у растений в условиях калийного голодания, нередко теряют всхожесть. Сильно снижается также устойчивость растений к различным заболеваниям и неблагоприятным климатическим условиям ( Баранов и Кореньков, 1956 г ).
в) физиологическая роль калия в растениях
Физиологическая роль калия в растениях изучена недостаточно и многие аспекты влияния калия на процессы, протекающие в растительном организме, остаются неясными. Можно и сейчас считать в основном правильной оценку положения, данную М. А. Егоровым еще в 1923 г.: тАЬНе только исчерпывающего, но и сколько-нибудь полного выяснения значения калия в жизни растения мы до сих пор не имеемтАЭ. Нак
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)