Строение и физиологические особенности рыб
Огловление
Форма тела и способы движения
Кожные покровы рыб
Мышечная система и электрические органы
Пищеварительная система
Дыхательная система и газообмен (New)
Кровеносная система
Нервная система и органы чувств
Железы внутренней секреции
Ядоносность и ядовитость рыб
Форма тела рыб и способы движения рыб
Форма тела должна обеспечивать рыбе возможность передвигаться в воде (среде, значительно более плотной, чем воздух) с наименьшей затратой энергии и со скоростью, соответствующей ее жизненным потребностям.
Форма тела, отвечающая этим требованиям, выработалась у рыб в результате эволюции: гладкое, без выступов тело, покрытое слизью, облегчает движение; шеи нет; заостренная голова с прижатыми жаберными крышками и сжатыми челюстями рассекает воду; система плавников определяет движение в нужном направлении. В соответствии с образом жизни выделено до 12 различных типов формы тела

Рис. 1 - сарган; 2 - скумбрия; 3 - лещ; 4 - рыба-луна; 5 - камбала; 6 - угорь; 7 - рыба-игла; 8 - сельдяной король; 9 - скат; 10 - рыба-еж; 11 - кузовок; 12 - макрурус.
Стреловидная - кости рыла вытянуты и заострены, тело рыбы по всей длине имеет одинаковую высоту, спинной плавник отнесен к хвостовому и располагается над анальным, чем создается имитация оперения стрелы. Эта форма типична для рыб, не перемещающихся на большие расстояния, держащихся в засаде и развивающих высокие скорости движения на короткий промежуток времени за счет толчка плавников при броске на добычу или уходе от хищника. Это щуки (Esox), сарганы (Belone) и др. Торпедовидная (ее часто называют веретеновидной) - характеризуется заостренной головой, закругленным, имеющим в поперечном разрезе форму овала телом, утонченным хвостовым стеблем, нередко с дополнительными плавничками. Она свойственна хорошим пловцам, способным к продолжительным перемещениям - тунцам, лососям, скумбриям, акулам и др. Эти рыбы способны в течение длительного времени плыть, если так можно выразиться, с крейсерской скоростью 18 км в час. Лососи способны совершать двух-трех метровые прыжки при преодолении препятствий во время нерестовых миграций. Максимальная скорость, которую может развить рыба, равна 100-130 км в час. Этот рекорд принадлежит рыбе-паруснику. Симметрично сжатое с боков тело - сильно сжато с боков, высокое при относительно небольшой длине и высокое. Это рыбы коралловых рифов - щетинкозубы (Chaetodon), зарослей донной растительности - скалярии (Pterophyllum). Такая форма тела помогает им легко маневрировать среди препятствий. Симметрично сжатую с боков форму тела имеют и некоторые пелагические рыбы, которым необходимо быстро менять положение в пространстве для дезориентации хищников. Такую же форму тела имеют рыба-луна (Mola mola L.) и лещ (Abramis brama L.). Несимметрично сжатое с боков тело - глаза смещены на одну сторону, что создает асимметрию тела. Она свойственна придонным малоподвижным рыбам отряда Камбалообразные, помогая им хорошо маскироваться на дне. В движении этих рыб большую роль играют волнообразные изгибания длинных спинного и анального плавников. Уплощенное в дорзовентральном направлении тело - сильно сжато в спинно-брюшном направлении, как правило, хорошо развиты грудные плавники. Такую форму тела имеют малоподвижные донные рыбы - большинство скатов (Batomorpha), морской черт (Lophius piscatorius L.). Уплощенное тело маскирует рыб в условиях дна, а расположенные сверху глаза помогают видеть добычу. Угревидная форма - тело рыб удлиненное, закругленное, имеющее вид овала на поперечном разрезе. Спинной и анальный плавники длинные, брюшных плавников нет, а хвостовой плавник небольшой. Она характерна для таких донных и придонных рыб, как угреобразные (Anguilliformes), передвигающихся, латерально изгибая тело. Лентовидная - тело рыб удлиненное, но в отличие от угревидной формы сильно сжато с боков, что обеспечивает большую удельную поверхность и позволяет рыбам обитать в толще воды. Характер движения у них такой же, как и у рыб угревидной формы. Такая форма тела характерна для рыбы-сабли (Trichiuridae), сельдяного короля (Regalecus). Макруровидная - тело рыбы высокое в передней части, суженное с задней, особенно в хвостовом отделе. Голова крупная, массивная, глаза большие. Свойственна глубоководным малоподвижным рыбам- макрурусообразным (Macrurus), химерообразным (Chimaeriformes). Астеролепидная (или кузовковидная) - тело заключено в костный панцирь, что обеспечивает защиту от хищников. Эта форма тела характерна для придонных обитателей, многие из которых встречаются в коралловых рифах, например для кузовков (Ostracion). Шаровидная форма свойственна некоторым видам из отряда Иглобрюхообразные (Tetraodontiformes) - рыбе-шару (Sphaeroides), рыбе-ежу (Diodon) и др. Эти рыбы плохие пловцы и передвигаются с помощью ундулирующих (волнообразных) движений плавников на небольшие расстояния. При опасности рыбы раздувают воздушные мешки кишечника, наполняя их водой или воздухом; при этом расправляются имеющиеся на теле шипы и колючки, защищающие их от хищников. Игловидная форма тела характерна для морских игл (Syngnathus). Их удлиненное, скрытое в костном панцире тело имитирует листья зостеры, в зарослях которой они обитают. Рыбы лишены боковой подвижности и перемещаются с помощью ундулирующего (волнообразного) действия спинного плавника.
Нередко встречаются рыбы, форма тела которых напоминает одновременно различные типы форм. Для ликвидации демаскирующей тени на брюхе рыбы, возникающей при освещении сверху, мелкие пелагические рыбы, например сельдевые (Clupeidae), чехонь (Pelecus cultratus (L.)], имеют заостренное, сжатое с боков брюшко с острым килем . У крупных подвижных пелагических хищников-скумбрий (Scomber), рыбы-меча (Xiphias gladius L.), тунцов (Thunnus)- киль обычно не развивается. Их способ защиты состоит в быстроте движения, а не в маскировке. У придонных рыб форма поперечного сечения приближается к равнобедренной трапеции, обращенной большим основанием вниз, что исключает появление тени на боках при освещении сверху. Поэтому большинство придонных рыб имеют широкое уплощенное тело.
КОЖА, ЧЕШУЯ И ОРГАНЫ СВЕЧЕНИЯ
В кожном покрове рыб различают два слоя: наружный слой эпителиальных клеток, или эпидермис, и внутренний слой из соединительнотканных клеток - собственно кожа, дерма, кориум, кутис.
Эпидермис защищает организм от проникновения в него веществ извне. Он состоит из многослойного эпителия, форма клеток и количество слоев которого варьируют у разных рыб. Но как правило самые верхние слои клеток плоские, по мере углубления они по форме приближаются к цилиндрическим. Эти клетки имеют ядро и способны размножаться. Новые слои клеток, отходящие от цилиндрических кнаружи, постепенно заменяют верхний отмирающий слой. Самый наружный слой эпителиальных клеток ороговевает, но в отличие от наземных позвоночных у рыб он не отмирает, сохраняя связь с живыми клетками. В течение жизни рыбы итенсивность ороговения эпидермиса не остается неизменной, наибольшей степени оно достигает у некоторых рыб перед нерестом: так, у самцов карповых и сиговых в некоторых местах тела (особенно на голове, жаберных крышках, боках и др.) появляется так называемая жемчужная сыпь - масса мелких белых бугорков, придающих коже шероховатость. После нереста она исчезает.
Между клетками эпидермиса имеются протоплазматические связи, по которым циркулирует лимфа, обеспечивающая питание клеток. В эпидермисе кожи рыб имеются различные железистые клетки, особенно слизевые. В целом кожа рыб отличается от кожи других позвоночных большим количеством слизи. Слизь образуется в специализированных железистых клетках, лежащих в эпидермисе.
Как правило, рыбы с хорошо развитым чешуйчатым покровом , выделяют слизи меньше (лососевые, окуневые). Рыбы, лишенные чешуи или чешуя которых редуцирована (круглоротые, некоторые сомы, линь, вьюн), выделяют слизи очень много. С бактерицидными свойствами слизи линя связывают повышенную устойчивость его к заражению паразитами, тогда как другие - представители семейства карповых такой устойчивостью не обладают. Роль слизи в жизненных отправлениях рыб очень велика: она уменьшает трение тела о воду (механическая защита), предотвращает попадание в организм паразитов и бактерий (бактерицидная защита), ускоряет свертывание крови в случаях ранений, способствует выведению веществ из организма, регулирует проникновение воды и солей (осмотическая регуляция), осаждает муть, склеивая частицы взвеси, в случае наличия в воде солей тяжелых металлов (медь, цинк, свинец, хром) коагулирующая слизь образует на коже защитный слой и, наконец, выделяет специфический видовой запах. Слизь некоторых рыб ядовита. Слизь миног вызывает у хищников нарушение пищеварения.
У некоторых рыб в эпидермисе имеются ядоотделительные железы, расположенные в основном у основания шипов или колючих лучей плавников. Иногда ядоотделительные клетки образуются и функционируют только во время размножения, в других случаях - постоянно. Уколы таких ядоносных рыб могут быть опасны для человека.
У ската - хвостокола ядоотделительные клетки расположены в эпидермисе у основания хвостового шипа. При уколе яд по желобку шипа поступает в рану и вызывает острую боль, сильный отек, озноб, тошноту и рвоту. В некоторых случаях наблюдается расстройство дыхания и даже смерть.
Особенно ядоносной является тихоокеанская бородавчатка. Многие лучи в ее плавниках снабжены ядоотделительными железами. При прикосновении к рыбе яд по бороздкам колючих лучей попадает в раны. Он разрушает красные кровяные тельца, парализует нервную систему и может оказаться смертельным не только для водных животных, но и для человека.
Ядоносных рыб следует отличать от ядовитых, у которых отдельные части тела (мышцы, кровь, икра, печень, кожа и др.) при употреблении в пищу вызывают отравление. Наибольшее число ядовитых рыб относится к семействам фахаков и кузовков, распространенных в водах Юго-Восточной Азии и тропической части Тихого океана.
Особенно ядовиты собаки-рыбы из отряда сростночелюстных. Наиболее опасным представителем этого отряда является фугу. В печени, икре, молоках и коже фугу содержится сильный яд. Мясо фугу съедобно и вкусно и ценится в некоторых странах. В Японии фугу разводят искусственно.
Среди акуловых и костистых (в большинстве глубоководных) рыб многие обладают способностью к люминесценции. Это свечение особого рода, при котором светоиспускание (в отличие от обычного - возникающего при тепловом излучении - основанного на тепловом возбуждении электронов и потому сопровождающегося выделением тепла) связано с генерацией холодного света (необходимая энергия образуется в результате химической реакции). Некоторые виды генерируют свет сами, другие своим свечением обязаны симбиотическим светящимся бактериям, которые находятся на поверхности тела или в специальных органах. В обоих случаях светящиеся клетки или органы располагаются в эпидермисе, но иногда во внутренних органах, например в кишечнике.
Нижний слой кожи - кориум - представляет собой проходящие в косом направлении волокна соединительной ткани. Здесь же находятся кровеносные сосуды и нервные окончания. В нижних слоях эпидермиса и в пограничных с ним слоях кориума залегают пигментные клетки - хроматофоры. Это звездчатые клетки со множеством отростков, включающие зернышки пигмента. Они определяют все разнообразие окраски рыб, особенно яркой в тропиках. Различие в окраске достигается сочетанием разных хроматофоров: меланофоры имеют зерна черного пигмента, ксантофоры - желтого, эритрофоры - красного; гуанофоры или иридоциты не имеют пигментных зерен, но содержат кристаллики гуанина, благодаря которым рыбы приобретают серебристую окраску.
Интенсивность окраски определяется состоянием хроматофоров: при их расширении пигментные зерна растекаются на большее пространство и окраска тела становится яркой. Если хроматофоры сокращаются, пигментные зерна скапливаются в центре, оставляя большую часть клетки неокрашенной и окраска тела бледнеет.
Форма пигментных клеток постоянна и у близких видов сходна, а состояние меняется в зависимости от различных факторов: температуры и газового режима водоема, возраста, пола, состояния организма (голод, размножение и т. п.), эмоций (возбуждение, страх) рыбы и др.
Сочетания разных цветов хроматофоров и отражательных поверхностей иридоцитов и их различные комбинации делают окраску рыб удивительно разнообразной. Сгущение и расхождение пигментов в клетках, а отсюда и изменение окраски тела рыб происходит под влиянием нервной системы и связано со зрительными восприятиями, причем центр просветления находится в продолговатом, центр потемнения - в промежуточном мозгу.
Изменение окраски в период размножения (появление брачного наряда), часто наблюдаемое в этот период различие в окраске самок и самцов происходят под влиянием гормональных факторов.
Разумеется, главное в расцветке большинства рыб - ее защитные свойства. Покровительственная окраска рыб, обитающих в поверхностных слоях воды, - темная спина и белое или серебристое брюшко - делает их малозаметными. Особенно удивительна маскировка у донных рыб - их окраска соответствует цвету дна. Наиболее удивительным мастером камуфляжа является обыкновенная камбала: она с легкостью хамелеона подделывается под камни, песок, темный ил.
Кожа подстилается рыхлой соединительнотканной прослойкой (подкожная соединительная ткань, подкожная клетчатка). У многих рыб в подкожной клетчатке откладывается жир.
В соединительнотканном слое кожи образуется чешуя. У рыб различают несколько видов чешуи: плакоидная, ганоидная и костная
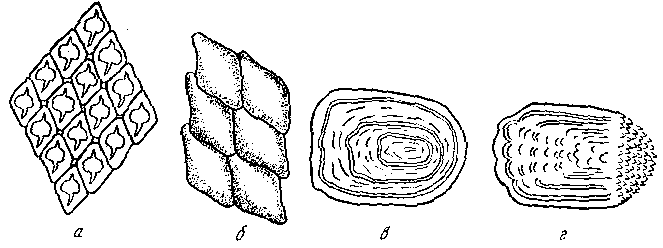
Рис. Форма чешуи рыб. а - плакоидная; б - ганоидная; в - циклоидная; г тАУ ктеноидная
Плакоидная - наиболее древняя, сохранилась у хрящевых рыб (акулы, скаты). Состоит из пластинки, на которой возвышается шипик. Старые чешуйки сбрасываются, на их месте возникают новые. Ганоидная - преимущественно у ископаемых рыб. Чешуйки имеют ромбическую форму, тесно сочленяются одна с другой, так что тело оказывается заключенным в панцирь. Чешуйки со временем не меняются. Названием своим чешуя обязана ганоину (дентинообразному веществу), толстым слоем лежащему на костной пластинке. Среди современных рыб ее имеют панцирные щуки и многоперы. Кроме того, она имеется у осетровых в виде пластинок на верхней лопасти хвостового плавника (фулькры) и жучек, разбросанных по телу (модификация нескольких слившихся ганоидных чешуек).
Постепенно видоизменяясь, чешуя теряла ганоин. У современных костистых рыб его уже нет, а чешуйки состоят из костных пластинок (костная чешуя). Эти чешуйки могут быть циклоидными - округлыми, с гладкими краями (карповые) и ктеноидными с зазубренным задним краем (окуневые). Обе формы родственны, но циклоидная как более примитивная встречатся у низкоорганизованных рыб. Бывают случаи, когда в пределах одного вида самцы имеют ктеноидную, а самки - циклоидную чешую (камбалы рода Liopsetta), или даже у одной особи встречаются чешуйки обеих форм.
Размеры и толщина чешуи у рыб сильно различаются- от микроскопических чешуек обыкновенного угря до очень крупных, величиной с ладонь чешуи трехметрового усача-тора, живущего в индийских реках. Лишь немногие рыбы не имеют чешуи. У некоторых она слилась в сплошной неподвижный панцирь, как у кузовка, или образовала ряды тесно соединенных костных пластинок, как у морских коньков.
Костные чешуйки, как и ганоидные, постоянны, не сменяются и лишь ежегодно увеличиваются в соответствии с ростом рыбы, и на них остаются отчетливые годичные и сезонные метки. Зимний слой имеет более частые и тонкие напластования, чем летний, поэтому он темнее летнего. По числу летних и зимних слоев на чешуе можно определить возраст некоторых рыб.
Под чешуей у многих рыб имеются серебристые кристаллики гуанина. Отмытые от чешуи, они являются ценным веществом для получения искусственного жемчуга. Из чешуи рыб изготовляют клей.
По бокам тела многих рыб можно наблюдать ряд выделяющихся чешуек с отверстиями, которые образуют боковую линию - один из важнейших органов чувств. Количество чешуи в боковой линии -
В одноклеточных железах кожи образуются феромоны - летучие (пахучие) вещества, выделяемые в окружающую среду и воздействующие на рецепторы других рыб. Они специфичны для разных видов, даже близкородственных; в некоторых случаях определена их внутривидовая дифференцировка (возрастная, половая).
У многих рыб, в том числе у карповых, образуется так называемое вещество страха (ихтиоптерин), которое выделяется в воду из тела пораненной особи и воспринимается ее сородичами как сигнал, извещающий об опасности.
Кожа рыб быстро регенерирует. Через нее происходит, с одной стороны, частичное выделение конечных продуктов обмена веществ, а с другой - поглощение некоторых веществ из внешней среды (кислород, угольная кислота, вода, сера, фосфор, кальций и другие элементы, играющие большую роль в жизнедеятельности). Большую роль играет кожа и как рецепторная поверхность: в ней располагаются термо-, баро-, хемо- и другие рецепторы.
В толще кориума образуются покровные кости черепа и пояса грудных плавников.
Через мышечные волокна миомеров, соединенные с ее внутренней поверхностью, кожа участвует в работе туловищно-хвостовой мускулатуры.
Мышечная система и электрические органы
Мышечную систему рыб, как и других позвоночных, разделяют на мышечную систему тела (соматическую) и внутренних органов (висцеральную).
В первой выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру.
Мышечная система взаимосвязана со скелетом (опора при сокращении) и нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определенным нервом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышц, которая в отличие от мышц млекопитающих невелика,
У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. Она обеспечивает плавание рыбы. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца - m. lateralis magnus) (рис. 1). Продольной соединительнотканной прослойкой эта мышца делится на спинную (верхнюю) и брюшную (нижнюю) части.
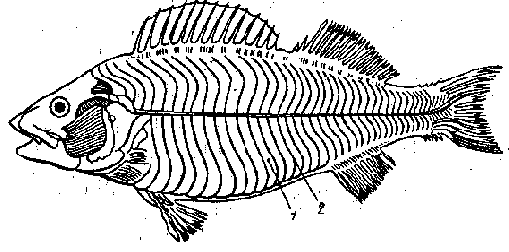
Рис. 1 Мускулатура костистой рыбы (по Кузнецову, Чернову, 1972):
1 - миомеры, 2 - миосепты
Боковые мышцы разделены миосептами на миомеры, число которых соответствует количеству позвонков. Наиболее отчетливо миомеры видны у личинок рыб, пока их тела прозрачны.
Мышцы правой и левой сторон, поочередно сокращаясь, сгибают хвостовой отдел тела и изменяют положение хвостового плавника, благодаря чему тело двигается вперед.
Над большой боковой мышцей вдоль тела между плечевым поясом и хвостом у осетровых и костистых лежит прямая боковая поверхностная мышца (m. rectus lateralis, m. lateralis superficialis). У лососевых в ней откладывается много жира. По нижней стороне тела тянется прямая брюшная мышца (m. rectus abdominalis); у некоторых рыб, например угрей, ее нет. Между ней и прямой боковой поверхностной мышцей располагаются косые мышцы (m. obliguus).
Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура), Плавники имеют свою мускулатуру.
Наибольшее скопление мускулов определяет и расположение центра тяжести тела: у большинства рыб он находится в спинной части.
Деятельность туловищных мышц регулируется спинным мозгом и мозжечком, а висцеральная мускулатура иннервируется периферической нервной системой, возбуждаемой непроизвольно.
Различают поперечнополосатые (действующие в значительной степени произвольно) и гладкие мышцы (которые действуют независимо от воли животного). К поперечно-полосатым относятся скелетные мышцы тела (туловищные) и мышцы сердца. Туловищные мышцы могут быстро и сильно сокращаться, однако скоро утомляются. Особенностью строения сердечных мышц является не параллельное расположение обособленных волокон, а разветвление их кончиков и переход из одного пучка в другой, что обусловливает непрерывную работу этого органа.
Гладкие мышцы также состоят из волокон, но гораздо более коротких и не обнаруживающих поперечной исчерченности. Это мышцы внутренних органов и стенок кровеносных сосудов, имеющие периферическую (симпатическую) иннервацию.
Поперечнополосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина - белка, легко связывающего кислород. Миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии.
Красные и белые волокна различны по целому ряду морфофизиологических характеристик: цвету, форме, механическим и биохимическим свойствам (интенсивность дыхания, содержание гликогена и т. д.).
Волокна красной мышцы (m. lateralis superficialis) - узкие, тонкие, интенсивно кровоснабжаемые, расположенные более поверхностно (у большинства видов под кожей, вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина;
в них обнаружены скопления жира и гликогена. Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых.
В мышце сердца (красной) мало гликогена и много ферментов аэробного обмена (окислительный обмен). Она характеризуется умеренной скоростью сокращений и утомляется медленнее, чем белые мышцы.
В широких, более толстых, светлых белых волокнах m. lateralis magnus миоглобина мало, меньше в них гликогена и дыхательных ферментов. Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко.
Красные мышцы постоянно деятельны. Они обеспечивают длительную и непрерывную работу органов, поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца.
При быстром движении, бросках активны белые мышцы, при медленном - красные. Поэтому наличие красных или белых волокон (мышц) зависит от подвижности рыб: "спринтеры" обладают почти исключительно белыми мышцами, у рыб, которым свойственны продолжительные миграции, кроме красных Боковых мышц имеются добавочные красные волокна в белых мышцах.
Основную массу мышечной ткани у рыб составляют белые мышцы. Например, у жереха, плотвы, чехони на их долю приходится 96,3; 95,2 и 94,9% соответственно.
Белые и красные мышцы различаются по химическому составу. В красных мышцах содержится больше жира, тогда как в белых мышцах больше влаги и белка.
Толщина (диаметр) мышечного волокна изменяется в зависимости от вида рыб, их возраста, величины, образа жизни, а у прудовых рыб - от условий содержания. Например, у карпа, выращенного на естественной пище, диаметр мышечного волокна составляет (мкм): у мальков - 5 .. 19, сеголетков - 14 .. 41, двухлетков - 25 .. 50.
Туловищная мускулатура образует основную долю мяса рыбы. Выход мяса в процентах общей массы тела (мясистость) неодинаков у разных видов, а у особей одного вида различается в зависимости от пола, условий содержания и др.
Мясо рыб усваивается быстрее, чем мясо теплокровных животных. Оно чаще бесцветно (судак) или имеет оттенки (оранжевый - у лососевых, желтоватый у осетровых и др.) в зависимости от наличия различных жиров и каротиноидов.
Основную массу белков мышц рыб составляют альбумины и глобулины (85%), всего же у разных рыб выделяют 4 .. 7 фракций белков.
Химический состав мяса (вода, жиры, белки, минеральные вещества) различен не только у разных видов, но и в разных частях тела. У рыб одного вида количество и химический состав мяса зависят от условий питания и физиологического состояния рыбы.
В нерестовый период, особенно у проходных рыб, расходуются резервные вещества, наблюдается истощение и, как следствие, уменьшается количество жира и ухудшается качество мяса. У кеты, например, во время подхода к нерестилищам относительная масса костей увеличивается в 1,5 раза, кожи - в 2,5 раза. Мышцы оводняются - содержание сухого вещества снижается более чем в два раза; из мышц практически исчезают жир и азотистые вещества - рыба теряет до 98,4% жира и 57% белка.
Особенности окружающей среды (в первую очередь пищи и воды) могут сильно изменять пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом. Качество мяса зависит и от диаметра мышечного волокна, а также количества жира в мышцах. В значительной мере оно определяется соотношением массы мышечной и соединительной тканей, по которому можно судить о содержании в мышцах полноценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки). Это соотношение изменяется в зависимости от физиологического состояния рыбы и факторов внешней среды. В мышечных белках костистых рыб на белки приходится: саркоплазмы 20 .. 30%, миофибрилл - 60 .. 70, стромы - около 2%.
Все многообразие движений тела обеспечивает работа мышечной системы. Она главным образом обеспечивает и выделение тепла и электричества в организме рыбы. Электрический ток образуется при проведении нервного импульса по нерву, при сокращении миофибрилл, раздражении светочувствительных клеток, механохеморецепторов и др.
Электрические органы
Своеобразно измененными мышцами являются электрические органы. Эти органы развиваются из зачатков поперечно-полосатой мускулатуры и расположены по бокам тела рыб. Они состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки (электроциты), переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя - положительно. Разряды происходят под действием импульсов продолговатого мозга. Вследствие разрядов вода разлагается на водород и кислород, поэтому, например, в заморных водоемах тропиков около электрических рыб скапливаются мелкие обитатели - моллюски, рачки, привлеченные более благоприятными условиями дыхания.
Электрические органы могут располагаться в разных частях тела: например, у ската морской лисицы - на хвосте, у электрического сома - на боках.
Генерируя электрический ток и воспринимая силовые линии,
искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния нескольких метров даже в мутной воде.
В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
1. Сильно электрические виды - имеют большие электрические органы, генерирующие разряды от 20 до 600 и даже 1000 В. Основное назначение разрядов - нападение и оборона (электрический угорь, электрический скат, электрический сом).
2. Слабоэлектрические виды - имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов - локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты).
3. Неэлектрические виды - не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10 .. 15 м в морской воде и до 2 м в пресной воде. Основное назначение генерируемого электричества - локация, ориентация, сигнализация (многие морские и пресноводные рыбы: например, ставрида, атерина, окунь и др.).
Пищеварительная система
В пищеварительном тракте настоящих рыб различают ротовую полость, глотку, пищевод, желудок, кишечник (тонкая, толстая, прямая кишка, заканчивающаяся анусом). У акул, скатов и некоторых других рыб перед анусом имеется клоака тАФ расширение, куда изливается прямая кишка и протоки мочевой и половой систем.
В ротовой полости рыб нет слюнных желез. Железистые клетки ротовой полости и глотки выделяют слизь, которая не имеет пищеварительных ферментов и способствует лишь проглатыванию пищи, а также защищает эпителий ротовой полости с вкрапленными вкусовыми почками (рецепторами).
Мощный и выдвижной язык имеют только круглоротые, у костистых рыб он не обладает собственной мускулатурой.
Рот обычно снабжен зубами. Наличием эмалевого колпачка и слоев дентина они напоминают зубы высших позвоночных. У хищников они располагаются как на челюстях, так и на других костях полости рта, иногда даже на языке; они острые. часто крючкообразные, наклонены внутрь к глотке и служат для схватывания и удержания жертвы. У многих мирных рыб (многие сельдевые, карповые и др.) на челюстях зубов нет.
Механизм питания координирован с дыхательным механизмом. Вода, всасываемая в рот при вдохе, несет и мелкие планктонные организмы, которые при выталкивании воды из жаберной полости (выдох) задерживаются в ней жаберными тычинками.
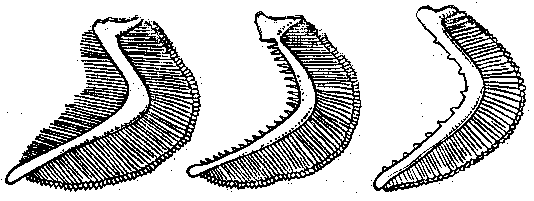
Рис. 1 Жаберные тычинки планктоноядной (а), бентосоядной (б), хищной (в) рыб.
Они так тонки, длинны и многочисленны у рыб, питающихся планктоном (планктонофагов), что образуют фильтрующий аппарат. Отцеженный комочек пищи направляется в пищевод. Хищные рыбы не нуждаются в отфильтровывании пищи, тычинки у них редкие, низкие, грубые, острые или крючковатые: они участвуют в удержании жертвы.
У некоторых бентосоядных рыб на задней жаберной дуге имеются широкие и массивные глоточные зубы, Они служат для перетирания пищи.
Следующий за глоткой пищевод, обычно короткий, широкий и прямой с сильными мускулистыми стенками проводит пищу в желудок. В стенках пищевода имеются многочисленные клетки, выделяющие слизь. У открытопузырных рыб в пищевод открывается проток плавательного пузыря.
Не все рыбы имеют желудок. К безжелудочным относятся карповые, многие бычки и некоторые другие.
В слизистой оболочке желудка имеются железистые клетки. вырабатывающие соляную кислоту и пепсин, расщепляющий белок в кислой среде, и слизь. Здесь у хищных рыб переваривается основная часть пищи.
В начальную часть кишечника (тонкую кишку) впадают желчный проток и проток поджелудочной железы. По ним в кишечник попадают желчь и ферменты поджелудочной железы, под действием которых происходит расщепление белков до аминокислот, жиров до глицерина и жирных кислот и расщепление полисахаридов до сахаров, главным образом глюкозы.
В кишечнике помимо расщепления питательных веществ происходит их всасывание, наиболее интенсивно протекающее в заднем участке. Этому способствует складчатое строение его стенок, наличие в них ворсинкообразных выростов, пронизанных капиллярами и лимфатическими сосудами, наличие клеток, секретирующих слизь.
У многих видов в начальной части кишечника помещаются слепые отростки тАФ пилорические придатки, число которых сильно варьирует: от 3 у окуня до 400 у лососей
Карповые, сомовые, щуки и некоторые другие рыбы пилорических придатков не имеют. При помощи пилорических придатков всасывающая поверхность кишечника увеличивается в несколько раз.
У рыб, не имеющих желудка, кишечный тракт представляет собой большей частью недифференцированную трубку, суживающуюся к концу. У некоторых рыб, в частности у карпа, передняя часть кишечника расширена и напоминает по форме желудок. Однако это лишь внешняя аналогия: здесь нет характерных для желудка желез, вырабатывающих пепсин.
Строение, форма и длина пищеварительного тракта разнообразны в связи с характером пищи (объектами питания, их усвояемостью), особенностями переваривания. Наблюдается определенная зависимость длины пищеварительного тракта от рода пищи. Так, относительная длина кишечника (отношение длины кишечника к длине тела.) составляет у растительноядных (пинагора и толстолобика)тАФб.. 15, у всеядных (карася и карпа) тАФ 2 .. 3, у хищных (щуки, судака, окуня) тАФ 0,6 .. 1,2.
Печень тАФ крупная пищеварительная железа, по размерам уступающая у взрослых рыб только гонадам. Ее масса составляет у акул 14.. 25%, у костистых тАФ 1 .. 8% массы тела Это сложная трубчато-сетчатая железа, по происхождению связанная с кишечником. У зародышей является его слепым выростом.
Желчные протоки проводят желчь в желчный пузырь (только у единичных видов его нет). Желчь благодаря щелочной реакции нейтрализует кислую реакцию желудочного сока. Она эмульгирует жиры, активирует липазу-тАФфермент поджелудочной железы.
Из пищеварительного тракта вся кровь медленно протекает через печень. В печеночных клетках кроме образования желчи происходит обезвреживание попавших с пищей чужеродных белков и ядов, откладывается гликоген, а у акул и тресковых (треска, налим и др.). тАФ жир и витамины. Пройдя через печень, кровь по печеночной вене направляется к сердцу.
Барьерная функция печени (очищение крови от вредных веществ) обусловливает ее важнейшую роль не только в пищеварении, но и кровообращении.
Поджелудочная железа тАФ сложная альвеолярная железа, также производная кишечника, является компактным органом только у акул и немногих других рыб. У большинства рыб она визуально не обнаруживается, так как диффузно внедрена в ткань печени (большей частью), и поэтому ее можно различить только на гистологических препаратах. Каждая долька связана с артерией, веной, нервным окончанием и протоком, выводящим секрет к желчному пузырю. Обе железы носят общее название hepatopancreas.
В поджелудочной железе вырабатываются пищеварительные ферменты, действующие на белки, жиры и углеводы (трипсин, эрепсин, энтерококиназа, липаза, амилаза, мальтаза), которые выводятся в кишечник.
У костистых рыб (впервые среди позвоночных) встречаются в паренхиме поджелудочной железы островки Лангерганса, в которых многочисленны клетки, синтезирующие инсулин, выделяемый прямо в кровь и регулирующий углеводный обмен.
Таким образом, поджелудочная железа является железой внешней и внутренней секреции.
Из мешкообразного впячивания спинной части начала кишечника образуется у рыб плавательный пузырь тАФ орган, свойственный только рыбам.
ДЫХАТЕЛЬНАЯ СИСТЕМА И ГАЗООБМЕН
Эволюция рыб привела к появлению жаберного аппарата, увеличению дыхательной поверхности жабр, а отклонение от основной линии развития тАФ к выработке приспособлений для использования кислорода воздуха. Большинство рыб дышит растворенным в воде кислородом, но есть виды, приспособившиеся частично и к воздушному дыханию (двоякодышащие, прыгун, змееголов и др.).
Основные органы дыхания. Основным органом извлечения кислорода из воды являются жабры.
Форма жабр разнообразна и зависит от видовой принадлежности и подвижности: мешочки со складочками (у рыбообразных), пластинки, лепестки, пучки слизистой, имеющие богатую сеть капилляров. Все эти приспособления направлены на создание наибольшей поверхности при наименьшем объеме.
У костистых рыб жаберный аппарат состоит из пяти жаберных дуг, располагающихся в жаберной полости и прикрытых жаберной крышкой. Четыре дуги на внешней выпуклой стороне имеют по два ряда жаберных лепестков, поддерживаемых опорными хрящами. Жаберные лепестки покрыты тонкими складками тАФ лепесточками. В них и происходит газообмен. Число лепестков варьирует; на 1 мм жаберного лепестка их приходится:
у щуки тАФ 15, камбалы тАФ 28, окуня тАФ 36. В результате полезная дыхательная поверхность жабр очень велика. К основанию жаберных лепестков подходит приносящая жаберная артерия, ее капилляры пронизывают лепесточки; из них окисленная (артериальная) кровь по выносящей жаберной артерии попадает в корень аорты. В капиллярах кровь течет в направлении, противоположном току воды
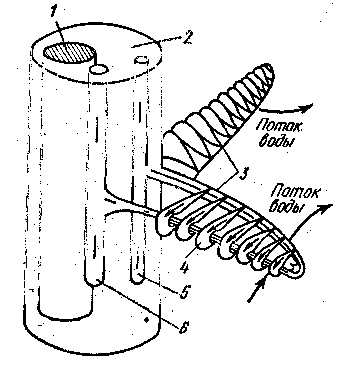
Рис.1 Схема противотока крови и воды в жабрах рыб:
1 - хрящевой стержень; 2 - жаберная дуга; 3 - жаберные лепестки; 4 - жаберные пластинки; 5 - приносящая артерия от брюшной аорты; 6 - выносящая артерия к спинной аорте.
Более активные рыбы имеют большую поверхность жабр: у окуня она почти в 2,5 раза больше, чем у камбалы. Противоток крови в капиллярах и омывающей жабры воды обеспечивает полное насыщение крови кисло
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)