Молекулярные основы наследственности
Содержание
Молекулярные основы наследственности
Хромосомы
Клеточный цикл
Мейоз и образование гамет
Строение хромосом
Наследование одиночных признаков
Независимая сегрегация и независимое комбинирование
Связь между генами и хромосомами
Рекомбинация
Связь между генами и белками
Гены и ДНК
Перенос генетической информации в клетке
Структура и сохранение геномной ДНК
Экспрессия и регуляция генов
Систематическое изучение наследственности начиналось со сложных в генетическом отношении объектов - растений и животных. Благодаря этим ранним исследованиям была сформулирована концепция неделимого гена как функциональной единицы наследственности и принято положение, что перенос генов от одного поколения к другому подвержен действию разных случайных факторов. Однако до понимания химической природы генов и механизма их функционирования было еще далеко. Исследование генетических молекул и тонких механизмов регуляции наследственности стало возможным лишь тогда, когда в качестве экспериментальных моделей начали использоваться бактерии и вирусы, о существовании которых первые генетики даже не подозревали. Только благодаря этим организмам впервые было показано, что дезоксирибонуклеиновая кислота, рибонуклеиновая кислота и белок - универсальные детерминанты генетического поведения. Стремительность дальнейшего прогресса в этой области и убедительность полученных результатов стали реальными благодаря особым биологическим свойствам микроорганизмов, которые позволяли проводить манипуляции, необходимые для анализа генетических структур. Аналогичные аналитические исследования более сложных генетических систем тогда были невозможны, поэтому на животных и растения этот прогресс не распространялся. Развитие технологии рекомбинантных ДНК разрушило труднопреодолимые технические и концептуальные барьеры на пути расшифровки и понимания сложных генетических систем. Неудивительно, что наши взгляды на структуру и функцию генов значительно изменились, а новое мышление в свою очередь радикально изменило перспективы биологии.
Некоторые предпосылки последних достижений можно обнаружить, изучая историю создания фундаментальных положений о наследственности и их последующих изменений. Основным препятствием на пути формирования единых принципов наследственности служило исключительное разнообразие живых форм. Первым, кто проследил аналогии между процессами воспроизведения животных и растений и ввел слова "самец" и "самка" применительно к участникам этого процесса, был ученик Аристотеля - Теофраст. Еще раньше греческие философы V в., воззрения которых оказали заметное влияние на последующее развитие научных идей, пришли к заключению, что, поскольку дети похожи на обоих родителей, оба пола вносят определенный вклад в формирование нового индивидуума. Они полагали, что этим вкладом является своего рода информация, сконцентрированная в мужском или женском "семени" и поступившая туда из разных частей тела зрелых индивидуумов. Демокрит, мнение которого не было общепринятым, предположил, что информация заключена в частицах, размер, форма и строение которых влияют на свойства потомства.
В начале XIX в., после создания более совершенных микроскопов, основной унифицирующей единицей в биологии стала клетка. Все организмы могли рассматриваться как одиночные, свободно живущие клетки или как сообщество клеток. Постоянное усовершенствование оптических систем микроскопа и новаторские методы подготовки и окрашивания материала позволяли все более детально описывать содержимое клеток не имеют ядра. Было установлено, что новые клетки появляются только в результате деления предсуществующих клеток.
В настоящее время все живые организмы подразделяют на две группы. Первая-эукариоты - многоклеточные организмы, клетки которых содержат оформленное ядро; внутри ядра заключены хромосомы-хранители генетической информации. Вторая - прокариоты - представлена одноклеточными бактериями, лишенными ядра, с хромосомами, находящимися в цитоплазме. За немногими исключениями, все клетки многоклеточного организма содержат одинаковый полный набор хромосом. Эукариотические организмы имеют более сложное строение и, как правило, содержат больше генетической информации. Кроме того, эукариоты способны к истинному половому воспроизведению и для многих из них этот способ обязателен для образования потомства. Одним из важных моментов процесса полового размножения является наличие в дочерних ядрах двух копий каждой хромосомы; такие эукариотические клетки называются диплоидными. Прокариоты, содержащие только одну хромосому, называются гаплоидами. При некоторых обстоятельствах у прокариот наблюдаются процессы, аналогичные по результату процессу оплодотворения у эукариот, вследствие которых они могут стать частично диплоидными; эти процессы широко используются в генетических исследованиях.
Сразу после принятия клеточной теории в изучении живых организмов выделились три направления: исследование хромосом, статистический анализ наследования одиночных признаков, выделение и характеристика компонентов хромосом.
В анафазе пары сестринских хроматид разделяются и каждый член пары движется по направлению к полюсу веретена. В это же время и нити веретена, и клетка начинают растягиваться. Когда в телофазе хроматиды достигают противоположных полюсов, вокруг каждого набора хроматид формируется новая ядерная оболочка и начинается деконденсация хромосом. Наконец, плазматическая мембрана разделяет два ядра и окружающую цитоплазму на две клетки. Хромосомы приобретают растянутую, диффузную форму, типичную для интерфазы, и процесс деления начинается снова.Б. Микрофотографии митоза в клетках лилии Haemanthus katherinae. Клетки окрашены иммунозолотом/серебром. Увеличение 600. правления параллельно развивались и превращались в важные научные дисциплины до момента их слияния в середине нашего века.
Во второй половине XIX в. продолжалось детальное изучение морфологии и поведения хромосом. Оказалось, что во всех клетках любого организма, за одним лишь существенным исключением, содержится одно и то же, вполне определенное число хромосом. Например, плодовая мушка Dro-sophila melanogaster имеет 8 хромосом, человек и летучая мышь-46, пшеница-20, носорог - 84. Хромосомы на основе сходства их морфологии могут быть разделены на гомологичные пары: 4 пары у D. melanogaster, 23 - у человека и т.д. Микроскопическое исследование фиксированных и окрашенных клеток дает лишь статическую картинку, но эти картинки можно расположить во временной последовательности, начиная с момента образования клетки при делении и кончая ее делением на две себе подобные. И тогда становится очевидным, что дупликация каждой хромосомы, происходящая в цикле клеточного деления, приводит к удвоению числа хромосом. При делении этот удвоенный набор распределяется таким образом, что каждая из двух дочерних клеток получает такое же число и тип хромосом, что и родительская клетка. Весь процесс в целом называется митозом.
События, происходящие в период от одного клеточного деления до другого, называются клеточным циклом. Фаза митоза цикла охватывает период деления и хромосом, и клеток. После расхождения клеток каждая дочерняя клетка вступает в период повышенной биосинтетической активности - в так называемую Gj-фазу. Gj-фаза заканчивается перед началом удвоения хромосом, или, в молекулярных терминах, с началом дупликации хромосомной ДНК; период репликации генома называется фазой синтеза. С момента завершения S-фазы в клетках инициируются события, характерные для митотической профазы,-части цикла, называемой Gj-фазой. В конце концов опять начинаются митоз и цитокинез, и цикл повторяется. Как правило, Gr, S - и G2-периоды, вместе составляющие интерфазу, занимают около 90% времени клеточного цикла, а М-фаза - менее 10%. Полное время прохождения клеточного цикла в клетках разного типа сильно варьирует в зависимости от условий роста. Основным показателем продолжительности всего цикла является продолжительность Gj-фазы. Например, покоя:
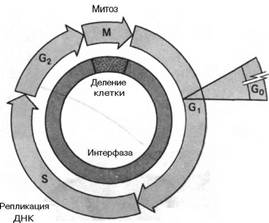
Клеточный цикл: митоз и цитокинез составляют М-фазу цикла, кульминацией которой является образование двух дочерних клеток. Каждая дочерняя клетка вступает в G1-период интерфазы и может начать новый клеточный цикл. За периодом G1 следует S-фаза, во время которой ДНК и хромосомы дуплицируются, и далее - фаза G2. Начало митоза означает конец интерфазы. Покоящиеся клетки задерживаются в фазе G1 и, как говорят, находятся в фазе G0. Обычно эукариотические клетки, которые не остановились в фазе G0, завершают цикл за 24 ч.
Мейоз: этапы деления диплоидной клетки на четыре гаплоидные дочерние клетки. Этот процесс отличается от митоза тем, что включает два клеточных деления и только один "раунд" репликации хромосом. На схеме показаны две пары гомологичных хромосом. Во время интерфазы хромосомы имеют вид тонких диффузных нитей. После репликации сестринские хроматиды остаются тесно связанными и начинают конденсироваться, что указывает на начало профазы. Затем гомологичные пары сестринских хро-матид приходят в тесное соприкосновение, образуя тетрады; этот процесс называется синапсисом. Начало мейотической метафазы I характеризуется дальнейшей конденсацией хромосом и дезинтеграцией ядерной мембраны. В анафазе I члены гомологичной пары сестринских хроматид начинают перемещаться к разным полюсам удлиняющейся клетки. К концу телофазы I и клеточного деления I образуются две дочерние клетки, в каждой из которых имеется по одной гомологичной паре сестринских хроматид. Второй раунд клеточного деления происходит без дополнительной дупликации хромосом и начинается с профазы II, с переходом в метафазу II. В стадии анафазы II две сестринские хроматиды, которые оставались до этого момента вместе, начинают перемещаться к противоположным концам удлиняющейся клетки. После телофазы II и клеточного деления II образуются четыре гаплоидные клетки - предшественники половых клеток. В каждую дочернюю клетку попадает только по одной хромосоме из исходных гомологичных пар.
|
|  |
Мейоз и образование гаметОбразование яйцеклеток и сперматозоидов подразумевает уменьшение нормального числа хромосом ровно вполовину; этот процесс называется мейозом. Гаметы, или половые клетки, гаплоидны, т.е. в них содержится по одному члену каждой пары гомологичных хромосом, и, таким образом, только половинное число хромосом каждого из родителей попадает во все другие, соматические, клетки организма потомка. Распределение хромосом в мейозе происходит случайно, поэтому любой из членов гомологичной пары может оказаться во вновь образовавшихся зародышевых клетках.
При оплодотворении гаплоидные наборы хромосом сперматозоидов и яйцеклеток объединяются. Таким образом восстанавливается полный набор гомологичных хромосомных пар, каждый из членов которых произошел из яйцеклетки и из сперматозоида соответствующих родителей. Диплоидное состояние оплодотворенной яйцеклетки поддерживается далее во всех соматических клетках механизмом митотического деления. Иногда зрелые организмы могут развиться из неоплодотворенных гаплоидных яйцеклеток или из оплодотворенных яйцеклеток с неполным набором родительских хромосом. Как уже отмечалось, любой из членов гомологичной пары может попасть в функциональную гамету. В зрелую яйцеклетку или сперматозоид попадает по одному члену каждой пары в процессе редукции числа хромосом в мейозе.
Легче всего наблюдать метафазные хромосомы. Под микроскопом их фотографируют или зарисовывают. В этой стадии хромосомы наиболее сконденсированны и образуют дискретные структуры. У многих организмов индивидуальные хромосомы и их гомологи легко различимы по размеру и форме. Каждая метафазная хромосома действительно состоит из двух идентичных частей, называемых сестринскими хроматидами, поскольку дупликация хромосомной ДНК протекает как раз перед метафазой, в S-фазе клеточного цикла.
У хромосомы имеется перетяжка, называемая центромерой. Положение центромеры для каждой хромосомы строго определено. С центромерой связаны специфические хромосомные функции; это последняя точка, соединяющая плечи сестринских хроматид перед полным расхождением при митотическом или II мейотическом делении. Сами плечи имеют вид
отдельных образований задолго до расхождения центромер в анафазе.
Образование гаплоидных гамет при мейозе и слияние двух гамет с образованием диплоидной клетки при оплодотворении. Обратите внимание на то, что у D. melanogaster, рассмотренной здесь в качестве примера, как и у других организмов, включая млекопитающих, две половые хромосомы у самца не гомологичны друг другу. При мейозе формируются два типа сперматозоидов, из которых один несет Х-, а другой - Y-хромосому. У самок, несущих пару Х-хромосом, в результате мейоза образуются гаметы одного типа. Пол потомков зависит от того, какую из хромосом - X или Y - несут оплодотворяющие сперматозоиды. У некоторых организмов негомологичную, определяющую пол хромосому несет самка.
Различие между областью центромеры и плечами хромосом становится очевидным после обработки определенными красителями. После окрашивания центромеры выглядят более плотными и компактными по сравнению с плечами. Такие плотные, интенсивно окрашиваемые хромосомные области называются гетерохроматиновыми. Гетерохроматин центромеры можно наблюдать после окрашивания даже в плохо различимых интерфазных хромосомах. Другие, негетерохроматиновые области хромосом принято называть эухроматиновыми. Эухроматиновые области окрашиваются гораздо менее интенсивно, чем гетерохроматиновые.
Концевые участки хромосом называются теломерами. Часто они тоже гетерохроматиновые. Нередко в митотических хромосомах можно наблюдать небольшие перетяжки, называемые районом ядрышкового организатора. В мейотических хромосомах они имеют вид утолщений. В пределах данного вида районы ядрышковых организаторов встречаются на одной или нескольких специфических хромосомах, и если они есть, то всегда находятся в одном и том же месте. В G1-фазе клеточного цикла некоторые ядрышковые организаторы начинают разрастаться; если их больше, чем один, то такие разросшиеся области объединяются в одну или несколько больших, почти сферических структур - нуклеолей. Этот рисунок достоверно воспроизводится, и каждую хромосому в наборе можно идентифицировать. На рис. I.9 представлен полный набор прометафазных хромосом в клетке человека. На этом изображении, называемом кариотипом человека, отражены относительный размер и форма хромосом наряду с положением центромеры и характерным видом полос.
В интерфазе хромосомы сильно растягиваются и, как правило, не видны. Встречаются, однако, и существенные исключения, которые уже много лет интенсивно исследуются. Секреторные клетки личинок некоторых насекомых разрастаются до огромных размеров и проходят несколько S-фаз без митоза и клеточного деления. В результате формируется комплекс из множества, иногда вплоть до тысячи, хроматид, которые остаются сцепленными и лежат рядом друг с другом, образуя толстые нити, называемые политенными хромосомами. Так же как и все интерфазные хромосомы, политенные хромосомы растянуты значительно сильнее, чем конденсированные метафазные хромосомы. При окрашивании политенных хромосом специальными красителями выявляется определенный рисунок чередования темных и светлых полос. В отличие от того, что наблюдается в высококонденсированных метафазных хромосомах, число полос огромно. Например, на четырех политенных хромосомах D. me-lanogaster можно насчитать почти 5000 темных полос, а в полном наборе из 23 метафазных хромосом человека видны по крайней мере 2000 полос.
Четко различимые морфологические признаки индивидуальных прометафазных и политенных хромосом стабильно воспроизводятся из поколения в поколение у данного вида. Необычная форма хромосом или характер полос наряду с атипичным числом хромосом сигнализируют о повреждении хромосомного материала. Наличие таких измененных хромосом часто связано с наследственными заболеваниями. Например, сегмент одной хромосомы иногда перемещается на совершенно неродственную хромосому, и такие перестройки сразу выявляются по необычному размеру или характеру полос. Подобные транслокации иногда бывают реципрокными, т.е. две неродственные хромосомы могут обменяться фрагментами. Другим примером изменений, или аберраций, хромосом служат делеции части нормальной хромосомы, дупликации некоторых областей и даже инверсии сегментов. Иногда наблюдаются потери хромосом или, напротив, появление лишних. Например, заболевание человека, известное как синдром Дауна, обусловлено присутствием трех копий 21-й хромосомы вместо обычных двух. Успехи в изучении структуры хромосом определялись выбором подходящих экспериментальных объектов. Так, огромные политенные хромосомы D. melanogaster стали излюбленной экспериментальной системой еще на заре развития области биологии, именуемой теперь цитогенетикой; систематическое изучение небольших по размеру хромосом человека и других млекопитающих могло начаться лишь с усовершенствованием экспериментальной техники в начале 50-х годов. Хромосомы прокариот не видны в световом микроскопе; недоступны для анализа с помощью светового микроскопа и мелкие, диффузные хромосомы таких низших эукариот, как дрожжи и трипаносомы.
Концепция гена восходит к началу 1860 г. и связана с именем Грегора Менделя, хотя до тех пор, пока другие ученые не повторили и не углубили его исследования в начале XX в., самого этого термина не существовало. Слово ген было введено В. Йогансеном в 1910 г. и относилось к гипотетической единице информации, регулирующей наследование индивидуальных признаков организма. Предположение о существовании генов было высказано на основании данных о статистическом распределении простых наследуемых признаков в потомстве известных родителей в течение нескольких поколений. В этих первых исследованиях генами оперировали как абстрактными статистическими понятиями, поскольку не было никакой информации относительно химической природы изучаемых признаков. Например, форма или цвет семян или цветков рассматривались как видимый наглядный наследуемый признак независимо от химической или метаболической основы этого свойства. Тем не менее, логический интеллектуальный фундамент, заложенный Менделем и его последователями, вполне соответствует нашим теперешним представлениям о химической структуре генов и тому, как эта структурная информация воплощается в свойства организма.
Взгляд Менделя на наследственность у эукариот определялся двумя главными обнаруженными им явлениями. Первое-существование независимой сегрегации. Любой организм содержит пару генов для любого одиночного наследуемого признака, при этом каждый из членов пары имеет либо отцовское, либо материнское происхождение. В каждом поколении члены каждой пары генов расходятся с образованием новых яйцеклеток или сперматозоидов, и во время оплодотворения формируются новые пары генов. Теперь члены пары называются аллелями и особенность признака зависит от объединения одинаковых аллелей или различных. Так, аллельные пары, детерминирующие некий признак, могут быть а1а1, а1а2 или а2а2 у разных индивидуумов. Тогда сперматозоиды или яйцеклетки должны содержать аллель а1 или а2. И хотя в каждом отдельном организме имеется не более двух разных аллелей определенного гена, в популяции данного вида циркулирует много различных аллелей. Например, могут существовать множественные формы гена а: а1, а2, а3, а4 и т.д., поэтому отдельные индивидуумы могут содержать такие пары, как а1а2, а2а2, а3а2, а1 а4, а4а5 и а4а4.
Второе важное наблюдение Менделя касалось независимого комбинирования различных аллельных пар генов, каждая из которых определяет различные признаки. Например, яйцеклетки или сперматозоиды в организме, содержащем аллельные пары а1а2 для признака а и Ь1Ь2 для b, могут иметь сочетания аллелей albl, alb2, a2bl или a2b2. При образовании гамет сегрегация аллельных пар "а" не зависит от сегрегации аллельных пар "b"; то же самое можно сказать и о других аллельных парах.
В начале XX в. была обнаружена корреляция между физическим поведением хромосом и положениями менделевской генетики. Каждый член аллельной пары генов мог быть ассоциирован с одной из хромосом пары, а независимое распределение аллелей можно было объяснить, если считать, что различные аллельные пары находятся на разных хромосомах. Томас Гент Морган и его коллеги доказали, что у D. melanogaster гены ассоциированы с хромосомами. Они выбрали этот организм для генетических исследований, поскольку короткое время генерации и большое число особей в потомстве, получаемом от каждого скрещивания, делает генетический анализ удобным и точным; кроме того, хромосомы D. melanogaster легкоразличимы в световом микроскопе. В ходе экспериментов было установлено, что наследование аллеля, приводящего к появлению у потомства белых, а не обычных красных глаз, всегда сцеплено с наследованием Х-хромосом и никогда-Y-хромосомы. Были обнаружены и другие аллели, коррелирующие с разными признаками, также связанные с наследованием Х-хромосом, и аллели, наследуемые совместно, сцепленными группами, но независимо от Х-хромосомы. Таким образом, стало очевидным, что число групп совместно наследуемых аллелей соответствует числу хромосомных пар. В ходе исследования было установлено, что аллели, ассоциируемые с различными хромосомами, распределяются в потомстве независимо, а группы аллелей, связанные с определенной хромосомой, остаются сцепленными и в потомстве.
Почти одновременно с выявлением групп сцепления были обнаружены и неожиданные исключения. Например, такие аллели, как а1 и b1 или а2 и b2, как правило, наследовались сцепленно, но иногда появлялись новые сочетания, a1b2 и а2^, которые наследовались в последующих поколениях. С помощью цитогенетического анализа было установлено, что при мейозе гомологичные хромосомы обвиваются друг вокруг друга, поэтому Морган предположил, что они могут обмениваться между собой частями, давая тем самым новые комбинации сцепленных аллелей. Этот процесс получил название кроссинговера или рекомбинации. Совершенно не зная химической природы этого явления, генетики использовали феномен рекомбинации в качестве основного инструмента генетических исследований. Определение частот рекомбинации между сцепленными парами аллелей у D. melanogaster позволило сделать три важных заключения: гены расположены в линейном порядке, и члены аллельных пар обычно занимают одинаковое относительное положение на гомологичных хромосомах; рекомбинация происходит только внутри одной группы сцепления; частота, с которой два разных сцепленных аллеля перекрещиваются, зависит от расстояния между ними на хромосоме. Относительное положение различных генов на хромосоме D. melanogaster, а в дальнейшем и других организмов было установлено именно исходя из этих принципов. К 1922 г. Морган и его коллеги смогли картировать несколько сотен генов на четырех хромосомах D. melanogaster.
Одно из первых предположений о том, как информация, заключенная в генах, проявляется в специфических свойствах клетки и целого организма, было высказано еще до того, как изобрели слово "ген". В первом десятилетии XX в. английский врач Арчибальд Гаррод заметил, что наследование некоторых метаболических идиосинкразий и других расстройств у людей происходит в соответствии с правилами Менделя. Он предположил, что причиной подобных наследственных расстройств служит недостаток или отсутствие особых ферментов, необходимых для нормального метаболизма.
Тогда же Гаррод высказал гипотезу, что детерминанты наследственности контролируют образование ферментов. Таким образом, способность к синтезу особых ферментов или даже их свойства связывались с генами. Предположение казалось очень заманчивым даже при отсутствии экспериментальных доказательств, потому что оно связывало имеющиеся в то время генетические данные, полученные для мух и растений, с биологией человека.
Дальнейшее развитие идеи Гаррода могли получить лишь с появлением новых экспериментальных подходов. В конце 1930-х годов такие подходы появились благодаря использованию в качестве экспериментальных объектов микроорганизмов. Вначале в центре внимания исследователей оказались низшие грибы из родов Aspergillus и Neurospora. Эти организмы хорошо росли в определенных условиях культивирования и достаточно быстро размножались. К середине 40-х годов было накоплено и проанализировано достаточно генетических и биохимических данных для того, чтобы прийти к выводу, что наличие или отсутствие фермента наследуемо и зависит от экспрессии одного гена. Джордж Бидл и Эдвард Татум обобщили связь между ферментом и геном в виде постулата один фермент - один ген. Поскольку ферменты - это белки, а многие белки состоят из более чем одного типа полипептидных цепей, постулат в дальнейшем стал формулироваться как "один полипептид - один ген". Проведенные исследования показали, что некоторые гены кодируют белки, не являющиеся ферментами, а другие гены контролируют образование молекул РНК, которые необходимы для синтеза белков.
Для обоих компонентов этой информационной цепочки часто используются такие термины, как генотип и фенотип, относящиеся соответственно к гену и признаку, который им кодируется. В общем виде генотип иногда трактуется как вся генетическая информация отдельной клетки или организма. Аналогично и термин фенотип применяется более широко для описания видимых свойств клетки или организма, будь то особые белки или функции либо морфологические и даже поведенческие признаки. Фенотип, как правило, является результатом взаимодействия между генетической информацией и условиями окружающей среды, в которой она реализуется. Термин геном применяется к совокупности хромосом, свойственных отдельному организму, в отличие от термина генотип, который относится к информации, заключенной в этих хромосомах.
К 1950 г. была обнаружена еще более заманчивая и многообещающая экспериментальная система для исследования связей между генами и функциями клетки. Обычная кишечная бактерия Escherichia coli имеет примитивные питательные потребности и делится каждые 20-60 мин, давая в потомстве огромное число клеток. У нее было обнаружено множество легко выявляемых генетически контролируемых физиологических признаков. Кроме того, использование мутантов, которые достаточно просто выделить и охарактеризовать, позволило идентифицировать гены, кодирующие специфические функции клетки. Таким образом был открыт путь для более формального генетического анализа и создания генетической карты единственной хромосомы E. coli. Еще одним преимуществом E. coli оказалось то, что эта бактерия является хозяином для нескольких вирусов, для которых в свою очередь характерно значительное генетическое разнообразие инфекционных свойств.
Бактериофаги, или, для краткости, фаги, оказались еще более удобной системой для генетических исследований. Два или даже больше фагов могут обмениваться фрагментами своих гомологичных геномов, порождая фаговое потомство с новыми генетическими свойствами. Фаговые геномы даже способны к обратимой интеграции с бактериальной хромосомой. При выщеплении из хромосомы фаг может включить в свой геном часть бактериального генома и, таким образом, стать носителем бактериальных генов. Анализ подобного обмена генетическим материалом показал, что даже такие примитивные организмы обладают упорядоченным геномом и индивидуальные гены могут составить генетическую карту.
Современная биохимическая генетика ведет свое начало от открытия ДНК в 1869 г. Фридрихом Мишером. Он установил, что вещество, экстрагируемое из гнойной массы и клеточных ядер, химически отличается от белков как по содержанию органического фосфора, так и по устойчивости к расщеплению протеолитическими ферментами. В течение последующих 85 лет были разработаны разные методы выделения ДНК с целью исследования природы ее химических составляющих и связи между ними. Кульминацией этих исследований стало установление основной структурной единицы ДНК: она состоит из фосфорилированного сахара, дезоксирибозофосфата, соединенного с азотистым основанием-либо с одним из пуринов, либо с одним из пиримидинов. Кроме того, с помощью биофизических методов было становлено, что молекула ДНК-это очень длинная цепочка, остов которой построен из дезоксирибозофосфатных единиц, соединенных друг с другом фосфодиэфирными мостиками; к каждой дезоксирибозной единице цепи присоединено либо пуриновое, либо пиримидиновое основание. В 1953 г. Джеймс Уотсон и Фрэнсис Крик обобщили накопленные к тому времени данные о составе и структуре ДНК, построив ставшую теперь классической теорию двойной спирали ДНК. Импульсом к ее созданию послужило обогатившее науку открытие Освальда Эвери и его коллег, а также Альфреда Херши и Маргарет Чейз, состоявшее в том, что только ДНК является носителем генетической информации. Центральная роль в наследственности, приписываемая хромосомам, могла быть теперь отнесена к ДНК, которую они содержат.
ДНК - не единственная нуклеиновая кислота, обнаруживаемая в клетке. Близкородственные молекулы - рибонуклеиновые кислоты - отличаются от ДНК в основном тем, что вместо дезоксирибозы содержат рибозу и чаще имеют одноцепочечную структуру.
Расшифровка структуры ДНК и установление ее центральной роли в наследственности увенчали накопленные наукой данные и позволили генетике из статистической и феноменологической науки превратиться в науку с преобладанием химических и молекулярных направлений развития. Незамедлительная бурная реакция ученых на открытие двойной спирали свидетельствовала об ее адекватности. Модель структуры ДНК не только соответствовала химическим и физическим данным, но и полностью отвечала функциям, присущим генетическому материалу. В линейной последовательности четырех пуринов и пиримидинов могло быть закодировано огромное количество информации, и в принципе эта структура могла обеспечить свою собственную репликацию. Расшифровка структуры ДНК проливала свет на самые разные аспекты биологии и создавала основу для объяснения многих разноречивых данных, полученных ранее. Она обеспечила фундаментальную целостность при интерпретации огромного многообразия жизненных форм. Раз и навсегда наследственность связывалась с определенной молекулярной структурой.
Проблемы механизмов переноса, перераспределения и экспрессии генетических признаков, долгое время не находившие решения, с начала 50-х годов перешли на молекулярный и химический уровни. Как реплицируются и рекомбинируют молекулы ДНК? Каким образом они сохраняются в последующих поколениях? Каким способом информация, закодированная в ДНК, обеспечивает образование фенотипических продуктов - белков? Как регулируется считывание информации, закодированной в ДНК, в процессе роста клеток или развития организма и при других физиологических состояниях? Как нарушаются эти процессы при заболеваниях? Эти и еще многие другие вопросы стояли в центре молекулярно-генетических исследований в течение последних 35 лет. Бурный прогресс в первые 20 из них был достигнут благодаря использованию систем прокариот и связан с идентификацией молекулярных структур, участвующих в процессах хранения, поддержания, передачи и использования генетической информации.
Информационные взаимоотношения между ДНК, РНК и белками теперь точно установлены. Репликация, с помощью которой создаются идентичные копии родительской молекулы ДНК, обеспечивает генетическую непрерывность в ряду поколений. Транскрипция ДНК с образованием РНК опосредует трансляцию этой информации на уровень белков. Итак, ДНК выполняет две основополагающие функции. Первая-это осуществление своей собственной репликации. Вторая - это формирование фенотипа через образование молекул РНК, участвующих в трансляции информации, содержащейся в ДНК, на язык белков. И, насколько это известно, только у эукариот информация может передаваться в обратном направлении, от РНК к ДНК, посредством процесса, именуемого обратной транскрипцией.
В основе переноса информации от ДНК к РНК или от РНК к ДНК лежит универсальная способность нуклеиновых кислот служить матрицей. Нуклеиновые кислоты направляют сборку идентичных или родственных молекул и непосредственно участвуют в процессе синтеза белка. Насколько известно, информация не передается от белков к нуклеиновым кислотам. Однако белки помимо самосборки осуществляют важнейшую функцию катализа и информационного переноса между нуклеиновыми кислотами.
Далее мы рассмотрим вкратце ключевые характеристики генетического аппарата и его функционирования: структурные особенности важнейших компонентов молекул - ДНК, РНК и белков - и то, как они работают, обеспечивая сохранение целостности генома и трансляцию генотипа организма в его фенотип. Эти вопросы детально рассматриваются в гл.1, 2 и 3, составляющих первую часть книги.
Все клеточные ДНК состоят из двух полинуклеотидных цепей, закрученных вокруг общей оси с образованием двойной спирали. Наружную поверхность спирали составляет остов каждой цепи, состоящий из повторяющихся остатков дезоксирибозы. Цепи удерживаются вместе благодаря водородным связям между пуриновыми основаниями одной цепи и пиримидиновыми - другой: аденин всегда спарен с тимином, а гуанин - с цитозином. В результате образования таких практически инвариантных пар последовательность оснований одной цепи однозначно определяет их последовательность в другой - иными словами, цепи двойной спирали ДНК комплиментарны.
Молекулы ДНК выполняют две разные функции. Первая - последовательность пуриновых и пиримидиновых оснований каждой цепи служит матрицей, с которой копируется новая цепь. Вторая - гены, составляющие ДНК, детерминируют синтез ферментов и других белков, необходимых для синтеза новых молекул ДНК. При репликации в особом участке двойной спирали ДНК происходит расплетание цепей. В результате каждая цепь начинает функционировать как матрица, на которой синтезируется новая, комплиментарная цепь. Таким образом, каждая из обеих образовавшихся дочерних спиралей получает одну цепь от родительской спирали, а другую - образованную в результате синтеза de novo. Несмотря на кажущуюся логическую простоту, процесс репликации в действительности очень сложен и для его осуществления необходимо множество белков. Важнейшими из них являются ферменты, называемые ДНК-полимеразами. Их роль в репликации состоит в сборке полинуклеотидных цепей из отдельных мононуклеотидов. Все ДНК-полимеразы удлиняют полинуклеотидную цепь последовательным добавлением отдельных дезоксинук
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)