Оптимальность как результат эволюции
1. Оптимальное пищедобывательное поведение
2. Явления компромисса в поведении животных
3. Функции затрат
4. Функциональные аспекты процесса принятия решения
5. Бюджет времени и бюджет энергии
Многие животные и растения кажутся идеально "сконструированными" для выполнения своих функций или достижения своих целей. Дарвин понимал, что возникновение столь явно совершенных адаптации можно объяснить с точки зрения естественного отбора. В настоящее время большинство биологов согласны с тем, что теория естественного отбора адекватно объясняет даже самые сложные адаптации, хотя некоторые исследователи все еще спорят с этим. Эта глава посвящена ряду проблем, которые возникают, если рассматривать поведение как результат эволюционного развития. Сначала мы обсудим закономерности оптимальной организации пищедобывательного поведения животного, т.е. тему, в которой весьма отчетливо выступают многие из указанных проблем, а затем рассмотрим вопрос о том, каким образом можно справиться с этими проблемами
Во времена Дарвина, чтобы опровергнуть теорию естественного отбора, часто прибегали к телеологическим аргументам. Ученые говорили, что с помощью такого пассивного и случайного механизма, каким представляется естественный отбор, нельзя объяснить очевидной целесообразности, присущей функциональной организации животных и растений. В своих аргументах телеологи использовали понятия цели или намерения для объяснения тех явлений, которые, казалось, невозможно было объяснить по-другому. В настоящее время телеологическую философию уже не считают пригодной при обсуждении проблем эволюции или закономерностей функционирования живого. Однако эта философия все еще играет значительную роль, когда речь идет о цели при обсуждении целенаправленного поведения отдельных организмов.
В последние годы в биологическом мышлении важную роль стало играть представление о запланированности, поскольку мы теперь понимаем, что естественный отбор - это формирующий агент. Современная эволюционная теория подразумевает, что животные стремятся приобрести такие свойства, которые гарантируют им, что в определенных стабильных условиях они не окажутся в худшем положении по сравнению с другими животными, с которыми они конкурируют. Иными словами, обитая достаточно долго в условиях стабильной среды, животные стремятся приобрести те свойства, которые в существующих условиях будут оптимальными. Это совсем не означает, что каждое животное прекрасно адаптировано ко всем условиям, поскольку все индивидуумы различаются между собой генетически и поскольку многих из них конкурирующие особи вытесняют из наиболее благоприятных местообитаний, к которым они были лучше всего приспособлены.
Многие животные способны выбирать из различных видов пищи ту, которая соответствует их физиологическим потребностям. Но для того чтобы получить пищу, животные должны расходовать энергию. Кроме того, им приходится тратить ценное время, расходовать такие физиологические "товары", как тепло и вода, а также подвергать себя опасности, рискуя быть съеденным хищником. Поэтому представляется очевидным, что способы добывания пищи неравноценны, если их рассматривать с точки зрения общей приспособленности животного.
Рассматривая процесс добывания пищи в этом аспекте, мы сначала должны выделить и охарактеризовать несколько основных эволюционных стратегий. Иногда полезно считать, что животные намеренно используют стратегию, направленную на увеличение будущего представительства своих генов, хотя по сути такая эволюционная стратегия является всего лишь пассивным результатом естественного отбора. Подобно тому, как мы можем говорить о стратегиях оборонительного и репродуктивного поведения, мы можем говорить и о стратегиях добывания пищи. Так, например, для некоторых видов животных характерна стратегия "сидеть и ждать", чтобы напасть на жертву из заранее выбранного укрытия. Одни виды охотятся, подкарауливая добычу, другие - преследуют ее. Приведем более конкретный пример. Весьма распространены случаи, когда для получения пищи один индивидуум извлекает пользу из пищедобывательных усилий другого индивидуума. Такие взаимоотношения могут наблюдаться как между двумя особями различных видов, так и между особями одного и того же вида. В данном случае такие взаимоотношения в принципе мало чем отличаются от некоторых стратегий полового поведения, когда самцы более низкого ранга спаривались с самками украдкой от доминантов.
Барнард и Сибли исследовали альтернативные стратегии при добывании пищи в группе живущих в неволе домовых воробьев. Оказалось, что в этом случае поставщики получают большую часть своей пищи за счет активного добывания мучных червей, предлагаемых экспериментатором. Что касается попрошаек, то они получают большую часть пищи, копируя поведение активно питающихся птиц или постоянно следуя за ними, вместо того чтобы самим активно разыскивать пищу. Иногда попрошайка выхватывал пищу у поставщика. Рассуждая теоретически, мы должны были бы ожидать, что "выигрыш" попрошаек должен увеличиваться с ростом числа поставщиков, поскольку увеличивается возможность паразитировать на них. Выигрыш поставщиков, напротив, должен уменьшаться с ростом числа попрошаек, поскольку у поставщиков становится меньше шансов извлечь выгоду из своих усилий, потраченных на добывание пищи.
Ситуация такого рода сама наталкивает на мысль проанализировать ее с позиций эволюционно стабильных стратегий, что и было сделано Барнардом и Сибли. Эти авторы обнаружили, что отдельные птицы не сменяют оппортунистически свою стратегию в соответствии со структурой стаи как целого. Попрошайки питаются лучше в том случае, когда поставщиков больше, однако при значительном превышении числа поставщиков питание попрошаек ухудшается. Вероятно, это происходит потому, что большое число поставщиков очень быстро уничтожает имеющуюся в наличии пищу.
Чтобы провести количественный анализ стратегии добывания пищи, необходимо допустить, что поведение животного предназначено для того, чтобы максимизировать некую величину. В том анализе, который провели Барнард и Сибли, в качестве выигрыша фигурировало число мучных червей, съеденных птицей за 10 мин. Иначе говоря, они предположили, что птицы максимизируют скорость потребления пищи. Совершенно очевидно, что такое допущение не вполне правомерно, поскольку можно было учесть и другие факторы, которые влияют на приспособленность животного в случае выбора той или иной стратегии. В частности, могут различаться "стоимости" этих двух стратегий. Однако, чтобы провести анализ такого типа, часто бывает удобно принять достаточно простой показатель приспособленности, например скорость получения пищи, или некую меру эффективности пищедобывания.
Когда хищник ловит свою жертву, ему приходится платить цену в виде времени, потраченного на то, чтобы поймать и съесть ее. Такое время часто называют временем "взаимодействия". Эта цена с лихвой окупается за счет чистой энергетической стоимости добычи, т.е. валовой ценности минус энергия, затраченная на поимку жертвы и ее переваривание. Рентабельность жертвы представляет собой чистую энергетическую стоимость, деленную на время взаимодействия. Когда животное имеет возможность выбирать добычу, то можно предположить, что оно выберет наиболее рентабельную жертву. Например, в одном исследовании ушастому окуню представлялась возможность охотиться в большом аквариуме на водяных блох - дафний. Исследователи обнаружили, что при низкой плотности жертвы рыбы не проявляют никакого предпочтения по отношению к мелким, средним или крупным дафниям. Однако при изобилии дафний рыбы выбирают из них самых крупных и наиболее рентабельных, игнорируя мелких. Этого результата можно было бы ожидать, если исходить из модели рентабельности, показанной на рис.1.
Правда, это можно объяснить и по-другому, а именно: рыба хватает любую дафнию, которая в данный момент кажется ей самой крупной. Ближайшая к ней дафния может показаться ей по размерам больше, чем более крупная, но находящаяся дальше от нее. По мере возрастания плотности дафний возрастает и вероятность появления рядом крупных дафний. Выбирая каждый раз добычу, которая выглядит крупнее, рыба может вести себя таким образом, как это предсказывает модель рентабельности. Заметим, однако, что эта модель отражает, что именно должно произойти в данной ситуации, а модель "брать самую большую" описывает правило, которым рыба может реально пользоваться.
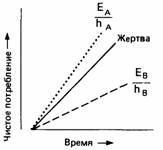
| Рис.1. Простая модель оптимального пищедобывательного поведения. Каждый тип жертвы характеризуется его рентабельностью E/h. Точками и штрихами вычерчены графики, крутизна которых отражает вероятные значения показателя для каждого типа жертвы. Если хищник поедает только более рентабельные жертвы, его поведение описывается сплошной линией. Ее крутизна меньше, чем крутизна пунктирной линии, поскольку хищнику приходится тратить время еще и на поиск более крупной жертвы. Если крутизна сплошной линии равна крутизне штриховой линии или меньше ее, то это означает, что для хищника имеет смысл поедать более мелкую добычу. Таким образом, по мере того как более крупные жертвы становятся все более редкими, сплошная линия поворачивается по часовой стрелке, и животное переключается на менее рентабельную добычу. |
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)