Особенности лихенофлоры сошненского лесничества Пинского района Брестской области
Проблема рационального использования природных ресурсов и их охраны может быть решена лишь комплексно совместными усилиями ученых и практиков на основе изучения и учета взаимосвязей и обусловленности многообразных природных процессов. Значительный интерес в этом плане представляют исследования в такой своеобразной группе растений, как лишайники: их видового состава, распространения, роли в различных биогеоценозах [1].
Лишайники относятся к сравнительно мало исследованным растительным организмам. Они представляют собой весьма своеобразную группу споровых растений, состоящую из двух компонентов тАУ гриба и одноклеточной, реже нитчатой водоросли, живущих совместно и образующих один целостный организм, обладающий иными, чем гриб и водоросль отдельно, морфологическими, анатомическими, физиологическими и экологическими свойствами. Известно более 20 000 видов и свыше 400 родов лишайников, встречающихся почти во всех природных зонах земного шара. Изучение их представляет теоретический и практический интерес.
Лишайники источник различных ценных медицинских препаратов и продуктов промышленного производства. Однако в настоящее время, в связи с недостаточностью их изученности, лишайники еще слабо используются.
Лишайники чутко реагируют на характер и состав субстрата, являясь хорошими индикаторами почвенно-грунтовых и микроклиматических условий, степени загрязненности воздуха в городах и населенных пунктах.
Лишайники могут быть использованы для датировки возраста различных предметов на основе измерения их слоевищ в диапазоне от нескольких десятилетий до нескольких тысячелетий [2].
Лишайники широко распространены в различных наземных биогеоценозах, но роль их в условиях Белоруссии особенно заметна в лесных сообществах. Здесь лишайники образуют группировки с большей биомассой, существенно влияют на жизнь леса [1].
Видовой состав лишайников очень разнообразен. Необходимость изучения лишайниковой флоры очевидна. В связи с этим, цель нашей работы на данном этапе тАУ изучить видовой состав лишайников различных экологических групп территории Сошненского лесничества, одного из фоновых лесничеств Пинского лесхоза, где подобные исследования вообще не проводились.
Задачи исследования:
1. Собрать лишайники с различных субстратов в разных фитоценозах;
2. Определить до вида и их систематизировать;
3. Проанализировать информацию о собранных лишайниках;
4. Составить гербарий наиболее часто встречаемых видов.
5. Полученную информацию передать в лесхоз.
1. ОБЗОР ЛИТЕРАТУРЫ
Лиша́йники (лат. Lichenes) тАУ симбиотические ассоциации грибов (микобионт) и микроскопических зелёных водорослей и/или цианобактерий (фотобионт); микобионт образует слоевище (таллом), внутри которого располагаются клетки фотобионта.
1.1 Этимология названия
Русское название лишайники получили за визуальное сходство с проявлениями некоторых кожных заболеваний, получивших общее название ВллишаиВ». Латинское название происходит от греческого λειχην (лат. Lichen) и переводится как бородавка, что связано с характерной формой плодовых тел некоторых представителей.
1.2 История исследования, систематическое положение
Usnea тАУ один из двух родов лишайников, описанных Теофрастом более двух тысяч лет назад. Первые описания известны из ВлИстории растенийВ» Теофраста, который указал два лишайника тАУ Usnea и Rocella, которую уже тогда использовали для получения красящих веществ. Теофраст предполагал, что они представляют собой наросты деревьев или водоросли. В XVII веке было известно только 28 видов.
Французский врач и ботаник Жозеф Питтон де Турнефор (рис. 1) в своей системе выделил лишайники в отдельную группу в составе мхов. Хотя к 1753 году было известно свыше 170 видов, Карл Линней описал только 80, охарактеризовав их как Влскудное крестьянство растительностиВ», и включил вместе с печёночниками в состав Влназемных водорослейВ».
Началом лихенологии (науки о лишайниках) принято считать 1803 год, когда ученик Карла Линнея Эрик Ахариус (рис. 2) опубликовал свой труд ВлMethodus, qua omnes detectos lichenes ad genera redigere tentavitВ» (ВлМетоды, с помощью которых каждый сможет определять лишайникиВ»). Он выделил их в самостоятельную группу и создал систему, основанную на строении плодовых тел, в которую вошли 906 описанных на то время видов.
Первым на симбиотическую природу в 1866 году на примере одного из видов указал врач и миколог Антон де Бари. В 1869 году ботаник Симон Швенденер (рис. 3) распространил эти представления на все виды. В том же году русские ботаники Андрей Сергеевич Фаминцин и Осип Васильевич Баранецкий обнаружили, что зеленые клетки в лишайнике тАУ одноклеточные водоросли. Эти открытия были восприняты современниками как ВлудивительнейшиеВ».
Сегодня лихенология является самостоятельной дисциплиной, смежной с микологией и ботаникой.
Традиционная систематика лишайников, оказывается во многом условна и отражает скорее особенности их строения и экологии, чем родственные отношения внутри группы, тем более что основывается она только на микобионте, а фотобионт сохраняет свою таксономическую самостоятельность.
Классифицируют лишайники по-разному, но в настоящее время рассматривают их как экологическую группу, уже не придавая им статуса таксона, поскольку независимость происхождения разных групп лишайников не вызывает сомнений, а группы, входящие в состав лишайников, помещают туда же, что и родственные микобионту грибы, не образующие лишайников. Для обозначения лишайников используют биноминальную номенклатуру, названия соотвествуют названию микобионта [3].

Рисунок 1 тАУ Жозеф Питтон де Турнефор тАУ впервые объединил лишайники в отдельную группу (в составе мхов).

Рисунок 2 тАУ Эрик Ахариус тАУ Влотец лихенологииВ», выделил лишайники в самостоятельную группу и впервые систематизировал их.

Рисунок 3 тАУ Симон Швенденер тАУ первооткрыватель симбиотической природы лишайников.
1.3 Микобионт, фотобионт и их симбиоз
Лишайники тАУ очень интересная и своеобразная группа низших растений.
В лишайнике сочетаются два организма с противоположными свойствами: водоросль (чаще зеленая), которая в процессе фотосинтеза создает органическое вещество, и гриб, потребляющий это вещество. Взаимоотношения их строятся следующим образом. Гриб получает от водоросли органические вещества тАУ углеводы, но в то же время как бы предоставляет водоросли, находящейся внутри тела лишайника, среду обитания, защиту от пересыхания и перегревания и т. д. Гриб снабжает водоросль достаточным количеством воды и растворенных в ней минеральных солей, которые он сам поглощает из окружающей среды (субстрата, атмосферного воздуха). Таким образом, хотя гриб в некоторой степени паразитирует на водоросли, но и она извлекает из совместной жизни с ним определенную пользу. Следовательно, в этом сожительстве наряду с паразитизмом имеются и черты симбиоза. В результате этих сложных взаимоотношений в процессе эволюции возник новый самостоятельный комплексный организм, имеющий новые, только ему присущие особенности строения и образа жизни и свои закономерности развития. При этом водоросль, выделенная из лишайника, в благоприятных условиях чаще всего может и вне его расти и развиваться самостоятельно, гриб же в этом случае обычно быстро погибает, так как в процессе приспособления к совместному существованию почти полностью потерял способность к самостоятельному развитию.
Поскольку основным формообразующим компонентом лишайников является гриб, то в последнее десятилетие среди ботаников получила распространение точка зрения, рассматривающая лишайники не как самостоятельную группу растений, а как грибы, включающие в свое тело водоросли, тАУ так называемые лихенизированные грибы, которые перешли к паразитизму на водорослях и в связи с этим значительно изменились. При таком подходе виды лишайников распределяются среди грибов в соответствии с типом плодового тела лишайникового гриба и особенностями его строения. Однако своеобразная форма тела лишайников, присутствие особых веществ, неизвестных ни у грибов, ни у водорослей, особенности обмена веществ и ряд других признаков позволяют рассматривать лишайники как самостоятельную группу организмов [4].
1.4 Строение лишайников
1.4.1 Внешнее строение таллома
По форме и величине лишайники разнообразны, их размеры тАУ от нескольких до десятков сантиметров. Вегетативное тело лишайников представлено слоевищем и в зависимости от образующегося в них пигмента может быть серым, сизым, зеленоватым, буро-коричневым, желтым, оранжевым или почти черным [5]. Пигменты лишайников откладываются в оболочках грибных гиф, реже в протоплазме. На интенсивность окраски сильное влияние оказывает освещенность: чем больше освещенность, тем ярче окраска.
Наиболее яркая окраска у лишайников высокогорных и приполярных районов Арктики и Антарктики. Это связано с тем, что пигменты, словно светофильтры, защищают лишайники от жесткого ультрафиолета, а в приполярных районах темноокрашенные лишайники быстрее нагреваются, снег и иней на их поверхности тает и превращается в капельножидкую влагу, необходимую лишайникам [6].
По внешнему виду различают три типа талломов лишайников: накипные (или корковые), листоватые и кустистые; эти типы связаны между собой переходными формами. Кроме них нередко различают еще чешуйчатый и филаментозный (нитевидный) типы талломов [7].
Таллом накипных лишайников представляет собой корочку, очень прочно срастающуюся с субстратом тАУ корой деревьев, обнаженной древесиной, поверхностью скал и камней. Этот таллом невозможно отделить от субстрата, на котором он растет, не повредив его. Поверхность такого накипного таллома может быть порошковатой, зернистой, бугорчатой или реже гладкой; окраска тАУ различной, обычно неяркой. У некоторых накипных лишайников почти весь таллом врастает в субстрат и на поверхности видны только плодовые тела лишайникового гриба; чаще это блюдцевидные апотеции.
Листоватые лишайники имеют вид чешуек или довольно крупных пластинок. Их таллом прикрепляется к субстрату обычно на большей своей части с помощью пучков грибных нитей (гиф) тАУ ризин или отдельных тонких гиф тАУ ризоидов. Лишь у немногих листоватых лишайников таллом срастается с субстратом только в одном месте с помощью мощного пучка грибных гиф, называемого гомфом. Между накипными и листоватыми лишайниками есть промежуточные формы, у которых таллом в центре накипной, а по краю листоватый, отстающий от субстрата.
У кустистых лишайников таллом состоит из ветвей или более толстых, часто ветвящихся стволиков. Такой кустистый лишайник срастается с субстратом только своим гомфом и растет вертикально либо наискось вверх (напочвенные виды) или свисая вниз (виды, растущие на стволах и ветвях деревьев). Между кустистыми и листоватыми талломами могут быть и переходные формы.
Для некоторых кустистых лишайников характерен так называемый первичный таллом, накипный или чаще листоватый, состоящий из мелких чешуек. На первичном талломе уже развивается вторичный, собственно кустистый, таллом в виде отдельных неразветвленных или разветвленных, вертикально стоящих веточек (рис. 4). Эти ветви вторичного таллома называются подециями и особенно характерны для рода кладония (С1аdоniа Hill. em. Vain.), у которого они очень разнообразны по форме. Подеции могут быть палочковидные, шиловидные, туповатые вверху или расширенные в виде бокальчика (сцифовидные), в виде кустиков с обильно разветвленными ветвями (рис. 4, 2). Сцифовидиые подеции часто пролиферируют, т. е. из центра сцифы или по ее краям вырастают новые сцифовидные подеции. У лишайников из родов кладония и стереокаулон (Stеrеосaulon Schreb.) на подециях развиваются чешуйки тАУ филлокладии. Они соответствуют по происхождению чешуйкам первичного таллома, как бы переходящим на подеции (рис. 4, 3). Первичный таллом может сохраняться в течение всей жизни лишайника, но у ряда видов он быстро исчезает и остаются только подеции.
Талломы большинства кустистых лишайников имеют радиальное строение (у кладонии, стереокаулона, уснеи, алектории). Есть кустистые лишайники с лентовидным ветвящимся талломом, где морфологически хорошо различаются две стороны тАУ верхняя и нижняя (так называемое дорзовентральное строение). Такой таллом характерен для рода цетрария (Сеtraria Ach.) [4].

Рисунок 4 тАУ Таллом кладоний: 1 тАУ таллом (а тАУ первичный, б тАУ вторичный, или подеций), 2 тАУ подеции кладоний (а, б тАУ шиловидные, или палочковидные, в тАУ с апотециями, г тАУ сцифовидный с пролиферацией по краю, е тАУ сцифовидный с пролиферацией в центре, ж тАУ кустистый), 3 тАУ талломы с филлокладиями, 4 тАУ филлокладии [а тАУ дланевидно(пальчато) разветвленные, б тАУ бородавчатые (бугорчатые), в тАУ коралловидно разветвленные, г тАУ щитовидно-чешуйчатые].
1.4.2 Грибы и водоросли, входящие в состав таллома лишайников
Среди лихенизированных грибов 90% связаны с зелеными водорослями, способными фиксировать атмосферный азот, а остальные 10% - с сине-зелеными водорослями из родов Nostoc Adanson, Scytonema Ag., Stigonema Ag., Ehichothrix Ag. и Calothrix Ag., усваивающими азот из атмосферы. Удивительно, что эти лишайники не характерны для местообитаний с недостатком азота, хотя водоросли в составе их таллома, безусловно, его фиксируют и высвобождают в значительном количестве [8].
Из одноклеточных зеленых водорослей в лишайниках встречаются виды родов хлорелла (Сhlоrеllа Beyer), цистококк (Сystoсоссиs Nüg); из нитчатых зеленых водорослей тАУ кладофора (С1аdорhоrа Kütz), плеврококк (Рleuroсоссus auct.), трептеполия (Тrеntероhliа Mart.). При этом нити плеврококка и трентеполии в талломе лишайника часто распадаются на отдельные клетки. Виды этих родов тАУ обычные, широко распространенные водоросли, часто встречающиеся как свободноживущие в пресных водоемах или па стволах деревьев. Нитчатая зеленая водоросль требуксия (Тrеbоихiа Puym.) живет только в талломах лишайников. Из сине-зеленых водорослей в талломах лишайников чаще всего встречаются представители рода носток (Nostoс Adanson); нитчатое тело этих водорослей в талломе лишайника также обычно распадается на отдельные участки. Водоросль, входящая в состав таллома лишайника, называется фотобионт.
Грибы, входящие в состав лишайников, в основном относятся к классу сумчатых (Аsсотусеtеs). Гифы гриба в подавляющем большинстве случаев (за исключением слизистых лишайников) составляют основу таллома лишайников, водоросль же как бы заключена между гифами гриба. Гриб, входящий в состав таллома лишайника, называется микобионт [4]
1.4.3 Анатомическое строение таллома
По анатомическому строению различают два типа талломов лишайников. В более примитивном, гомеомерном талломе клетки или нити водоросли более или менее равномерно распределены между гифами гриба по всей толщине таллома. Такой таллом имеют слизистые лишайники, содержащие в качестве фикобионта нитчатые сине-зеленые водоросли. Нити этих водорослей окружены большим количеством слизи, в которой по всем направлениям проходят грибные гифы, что хорошо видно на поперечном срезе таллома под микроскопом или через 10-кратную лупу. В сухом состоянии такие талломы имеют вид буроватых или черноватых морщинистых корочек. Во влажную погоду они быстро впитывают влагу, разбухают и приобретают форму листоватых лишайников [9]. Один из таких лишайников тАУ коллема (Соllета Web.), виды которого широко распространены на скалах и камнях на Черноморском побережье Крыма, Кавказа и сопредельных областей. Это черноватые или темно-оливковые подушечки, образуемые извилистыми приподнятыми лопастями.
Более сложно построен гетеромерный таллом. На поперечном срезе такого таллома под микроскопом хорошо различимы элементы его строения. Верхняя кора таллома образована плотным переплетением гиф гриба. Дальше идет гонидиальный слой, состоящий из клеток водоросли. Гифы гриба, заходящие в гонидиальный слой, образуют мелкие разветвления, которые плотно примыкают к клеткам водоросли. Здесь гриб получает от фотосинтезирующей водоросли углеводы. Дальше расположена сердцевина (слой из рыхлопереплетенпых гиф гриба), с помощью, которой внутри таллома поддерживаются определенная влажность и воздушная среда, необходимая и для самих гиф, и для клеток водоросли. За сердцевиной расположена нижняя кора из плотно переплетенных гиф гриба. Гетеромерный таллом хорошо выражен у листоватых лишайников и у кустистых лишайников с лентовидным дорзовентральным талломом.
У кустистых лишайников с цилиндрическим радиальным талломом (сцифовидный, палочковидный и т. д.) таллом имеет гетеромерно-радиальное строение. Под корой, одевающей снаружи ветви такого таллома, лежит гонидиальный слой, идущий вокруг всего таллома, а внутрь от него расположена сердцевина. У накипных лишайников с гетеромерным талломом никогда не бывает нижней коры, они срастаются с субстратом сердцевинными гифами.
Развитие и выраженность слоев гетеромерного таллома у разных видов лишайников различны. У ряда листоватых и кустистых лишайников с гетеромерным талломом происходит местный разрыв корового слоя и возникают плоские, неотчетливо очерченные пятна, обычно более светлые, чем сама кора (макулы, или цифеллы); они служат для проведения воздуха в сердцевину таллома и у ряда видов являются определительным систематическим признаком. Встречаются макулы у рода цетрария, например у Влисландского мхаВ» (Сеtraria islandicа L.)
1.4.4 Органы спороношения лишайникового гриба
На талломе лишайника из грибных гиф формируются плодовые тела гриба со спорами. Это в основном расположенные на поверхности таллома апотеций или погруженные в таллом кувшиновидные перитеции.
В апотециях и перитециях формируются споры для размножения лишайникового гриба.
Перитеции встречаются у небольшого числа лишайников. Апотеции образуются у многих видов лишайников. Они имеют чаще блюдцевидную форму, но встречаются и более или менее выпуклые или почти шаровидные. Самые крупные апотеции имеют диаметр более 1 см, но лишайников с такими апотециями немного. У громадного большинства лишайников диаметр апотециев от одного до нескольких миллиметров. Они разбросаны по поверхности листоватого таллома, чаще в его середине, или расположены по краям его лопастей. У кустистых лишайников апотеции обычно находятся на концах веточек или сциф. Они бывают сидячими или реже на небольших ножках приподнимаются над талломом. Апотеции могут быть окрашены в один цвет с талломом, или поверхность апотеция (так называемый диск апотеция) имеет другую окраску.
По строению различают три типа апотециев: леканориновый, лецидеиновый и биаториновый.
Леканориновый апотеций по своему анатомическому строению сходен с талломом лишайника. Его диск имеет талломный (слоевищный) край, образованный талломом лишайника и состоящий из гиф гриба и клеток водоросли. Этот край окрашен так же, как и таллом лишайника, и отличается по окраске от самого диска. В самом талломе под леканориновый апотецием также имеются клетки водоросли (рис. 5, 2).
Лецидеиновый апотеций имеет край, состоящий только из гиф гриба и окрашенный в тот же цвет, что и диск апотеция. В этом крае, а также в талломе под лецидеиновым апотецием клеток водорослей нет. Лецидеиновые апотеции имеют твердую консистенцию и обычно темную окраску (рис. 5, 3).
Биаториновый апотеций имеет такое же строение, как лецидеиновый, но отличается яркой окраской и мягкой консистенцией.
1.5 Размножение лишайников
Споры лишайникового гриба, выбрасываемые из перитециев и апотециев, прорастают в гифы, которые, переплетаясь, образуют зачаточный таллом лишайника, или прототаллом. Чтобы в дальнейшем из него возник настоящий лишайник, необходим контакт прототаллома с определенным видом водоросли, а также определенные внешние условия. В противном случае прототаллом быстро погибает. Таким образом, споры гриба, образующиеся в апотециях и перитециях, большого значения для размножения лишайников не имеют, поскольку сочетание необходимых для этого условий в природе встречается редко.
Более важны для размножения лишайников такие образования, в которых одновременно присутствуют гифы гриба и клетки водоросли. Это соредии и изидии. Они служат для размножения лишайника как целого организма. Попав в благоприятные условия, они дают начало непосредственно новому таллому. Соредии и изидии встречаются чаще у листоватых и кустистых лишайников.
Соредии представляют собой мельчайшие образования в виде пылинок, состоящих из одной или нескольких клеток водоросли, окруженных гифами гриба (рис. 5, 4). Формирование их обычно начинается в гонидиальном слое. Вследствие массового образования соредии количество их увеличивается, они давят на верхнюю кору, разрывают ее и оказываются на поверхности таллома, откуда легко сдуваются при любом движении воздуха или смываются водой. Скопления соредии называют соралями. Наличие и отсутствие соредии и соралей, их расположение, форма и окраска постоянны для определенных лишайников и служат определительным признаком.
Иногда при отмирании лишайников их таллом превращается в порошковатую массу, состоящую из соредии. Это так называемые лепрозные формы лишайников (от греческого слова ВллепросВ» тАФ ВлшероховатыйВ», ВлнеровныйВ»). В этом случае определить лишайник почти не представляется возможным.
Соредии, разносимые ветром и дождевой водой, попав в благоприятные условия, постепенно образуют новый таллом. Возобновление нового таллома из соредии происходит очень медленно. Так, у видов из рода кладония нормальные чешуйки первичного таллома развиваются из соредии только через срок от 9 до 24 месяцев. А для развития вторичного таллома с апотециями требуется от одного до восьми лет в зависимости от вида лишайника и внешних условий.
Если условия для развития нового таллома не совсем благоприятны, но все же возможен рост грибных гиф и размножение водорослей, то из соредии развиваются новые соредии и возникает описанная выше лепрозная форма, имеющая вид порошковатого пятна.
Изидии встречаются у меньшего числа видов лишайников, нежели соредии и сорали. Они представляют собой простые или коралловидно разветвленные выросты, обычно густо покрывающие верхнюю сторону таллома. В отличие от соралей изидии снаружи покрыты корой, часто более темной, чем таллом. Внутри, под корой, они содержат водоросли и грибные гифы (рис. 5, 5). Изидии легко отламываются от поверхности таллома. Обламываясь и распространяясь с помощью дождя и ветра, они, так же как и соредии, могут при благоприятных условиях образовывать новые талломы лишайников.
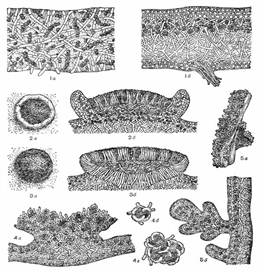
Рисунок 5 тАУ 1 тАУ разрез таллома (а тАУ гомеомерного, б тАУ гетеромерного) 2 тАУ леканориновый апотеций (а тАУ вид сверху, б тАУ разрез), 3 тАУ лецидеиновый апотеций (а тАУ вид сверху, б тАУ разрез), 4 тАУ строение соралей (а тАУ сораль, б, в тАУ отдельные соредии), 5 тАУ изидии (а тАУ внешний вид, б тАУ разрез).
Многие лишайники не образуют апотециев, соредий и изидии и размножаются участками таллома, которые легко отламываются от хрупких в сухую погоду лишайников ветром или животными и ими же переносятся. Особенно широко распространено размножение лишайников участками таллома в арктических областях. В наибольшей степени размножение кусочками таллома присуще накипным эпилитным лишайникам. Появление трещин в скалах и камнях, покрытых лишайниками, а также появление трещин в самом талломе накипного лишайника из-за неодинакового отношения к нагреванию и увлажнению камня и самого лишайника ведет к разламыванию таллома и отделению небольших его участков. Эти отделившиеся кусочки таллома и переносятся ветром и водой на новые места, давая начало новым талломам.
Для некоторых лишайников (около 100 видов из разных родов) характерно наличие на поверхности, а иногда и внутри таллома образований разного размера в виде разбросанных небольших вздутий, бородавочек, булавовидных, шаровидных и коралловидных выростов. Это цефалодии. Они могут покрывать верхнюю или нижнюю сторону листоватых талломов или развиваться по бокам подециев либо веточек кустистых талломов. В отличие от изидии они более разнообразной формы и разбросаны более редко. На срезе под микроскопом видно, что они имеют гомеомерное строение и состоят из беспорядочного скопления гиф, среди которых равномерно распределены клетки или нити сине-зеленых водорослей. Реже цефалодии имеет гетеромерное строение. Водоросли в цефалодиях всегда сине-зеленые, тогда как в талломе лишайника, несущего цефалодии, водоросли всегда зеленые. Образование цефалодиев обусловлено проникновением в таллом не свойственных данному виду лишайника сине-зеленых водорослей, которые вызывают местное разрастание гиф гриба, ВлобрастающегоВ» эти водоросли. Роль цефалодиев для самого лишайника не совсем ясна. Предполагается и отчасти доказано экспериментально, что они увеличивают питание лишайника, живущего на бедном субстрате. Сине-зеленые водоросли цефалодия способны фиксировать атмосферный азот, который затем в виде азотистых соединений усваивается всем талломом лишайника тАУ составляющими его грибом и зеленой водорослью, которые сами не могут фиксировать атмосферный азот.
Цефалодии специфичны для ряда видов и имеют значение определительного (систематического) признака. Характерный пример лишайника с цефалодиями тАУ напочвенный вид пельтигера пупырчатая (Peltigeraaphtnosa(L.) Willd.).
1.6 Распространение и экологические группы лишайников
Лишайники насчитывают около 20 тысяч видов и широко распространены по земному шару тАУ от полярных холодных скал до раскаленных камней пустынь.
Закономерности их географического распространения изучены еще недостаточно. С одной стороны, отмечается приуроченность определенных видов к тем или иным природным зонам, (например, есть виды, приуроченные к зоне таежных лесов, такие, как уснея длиннейшая тАУ Usnea longissima Ach.). Но есть виды, которые в своем распространении связаны не столько с природными условиями определенной зоны, сколько с условиями, которые повторяются в нескольких природных зонах [виды, связанные с океаническим климатом, многие арктоальпийские виды и т. д.; стереокаулон альпийский (Stereocaulon alpinum Laur.), например, растет на Крайнем Севере на каменистой почве тундр, а также в альпийском поясе различных горных массивов (Алтай, Кавказ)]. Географическое распространение многих лишайников прямо связано с их выборочным отношением к субстрату, хотя последний и не является основной причиной, ограничивающей их распространение. Безразличные к субстратам виды лишайников имеют более широкий ареал (область распространения), чем виды, более требовательные к определенному субстрату. Есть лишайники космополиты, распространенные по всему земному шару, но таких относительно немного.
По приуроченности к субстрату лишайники подразделяют на несколько экологических групп.
Напочвенные, или эпигейные, лишайники. Виды этой группы должны выдерживать сильную конкуренцию со стороны быстрорастущих высших растений, особенно травянистых. Поэтому они редко встречаются на плодородных почвах и достигают большего развития в местах, мало пригодных для высших растений из-за незначительной питательности субстрата или неблагоприятных климатических условий, например на песчаных почвах, в тундрах, полупустынях, на торфяниках и т. д.
Напочвенные лишайники могут расти как на открытых местах, так и в лесах. Лишайники открытых пространств произрастают вдоль дорог, на старых пожарищах, на бедных сухих и болотистых лугах, на опушках. Это пельтигера собачья (Peltigera canina (L.) Willd.), пельтигера бородавчатая (P. aphthosa (L.) Willd.), пельтигера прорывающаяся (P. erumpens (L.) Willd.), Влисландский мохВ» (Cetraria islandica (L.) Ach.), стереокаулон войлочный (Stereocaulon tomentosum Fr.), различные виды рода кладония (Cladonia Hill. em. Vain). Особенно пышно разрастаются напочвенные лишайники открытых пространств в тундре и лесотундре, где часто занимают громадные площади и определяют характер ландшафта. Это широко распространенные виды Влоленьего мхаВ»: кладония оленья (C. rangiferina (L.)), кладония приальпийская (C. alpestris (L.) Rabh.), кладония лесная (C. sylvatica f. tenius). Кроме них здесь растут различные кустистые виды рода цетрария (Cetraria Ach.), виды родов алектория (Alectoria Ach.), стереокаулон и др. Очень характерна для тундры охролехия двуполая (Ochrolechia androgyna (Hoffm.) Arn.), иногда на больших площадях образующая, беловатую корочку на мхах, некоторых цветковых растениях и даже на других лишайниках. Достигая большого развития, охролехия препятствует фотосинтезу и как бы удушает растения, на которых поселилась. Широко распространены в тундре также охролехия виннокаменная (O. tartarea (L.) Massal.) и охролехия холодная (O. frigid (Sw.) Lynge).
Напочвенные лишайники открытых пространств встречаются также в сухих степях и полупустынях, на скалах и каменных россыпях в высокогорных районах. Они образуют две четкие подгруппы видов. Первая подгруппа тАУ ВлкочующиеВ» лишайники, не срастающиеся с почвой и переносимые ветром с места на место, как перекати-поле. Это в основном мелколистоватые лишайники: аспицилия съедобная, или Вллишайниковая маннаВ» (Aspicilia esculenta (Pall.) Flag.), пармелия блуждающая (Parmelia vagans Nyl.), пармелия грубоморщинистая (P. ryssolea (Ach.) Nyl.), корникулярия степная (C. steppae Ach.). Вторая подгруппа тАУ прикрепленные к почве лишайники; в основном это накипные виды, образующие в сухую погоду на почве малозаметные налеты или корочки. Среди них есть виды, приуроченные к песчаным [рамалина многообразная (Ramalina polymorpha (L.) Ach.), рамалина скрученная (R. strepsilis (L.) Ach.), пармелия темно-бурая (Parmelia pulla (L.) Ach.) и др.], известковым [виды родов веррукария (Verrucaria Wigg. em. Th. Fr.) и коллема (Collema Web.)] и глинистым [беомицес розовый (Baeomyces roseus Pers.) и беомицес рыжий (Baeomyces rufus (Huds.))] почвам.
Напочвенные лишайники в лесу, особенно негустом, представлены различными видами кладоний, пельтигер. Особенно большого развития лишайниковый покров достигает в сухих сосняках. Это так называемые боры-беломошники. Покров из кладоний бывает сильно развит и в лиственничных лесах Восточной Сибири.
К представителям напочвенных лишайников лесов близко примыкают по своей экологии лишайники, развивающиеся на гниющих пнях и на основаниях стволов деревьев. В большинстве своем они теневыносливы и влаголюбивы. Это цетрария сосновая (Cetraria pinastri (Scop.) Röhl.), кладония пальчатая (Cladonia digitata Schaer.), упомянутые уже виды кладоний, составляющие Влолений мохВ», и т. д. Эти виды являются переходными к следующей экологической группе.
Эпифитные лишайники поселяются на деревьях и кустарниках. Среди них также можно выделить несколько подгрупп: эпифилльные лишайники, растущие на листьях деревьев и кустарников; настоящие эпифитные лишайники, растущие на коре; эпиксильные лишайники, растущие на обнаженной и обработанной древесине.
Эпифилльные лишайники сравнительно немногочисленны. Распространены они главным образом в тропиках и отчасти в субтропиках на старых листьях вечнозеленых растений. Большей частью они используют зеленый лист только как опорный субстрат. Эпифилльные лишайники живут в основном на поверхности листа, не проникая внутрь него и не причиняя ему вреда. Но есть виды, которые проникают под кутикулу и прикрепляются к наружным стенкам клеток эпидермиса листа. Эпидермис от этого или страдает мало, или отмирает, и в этом случае на листьях обычно образуются серые или серо-зеленые пятна. На территории бывшего СССР немногочисленные эпифилльные лишайники встречаются в основном на юге Черноморского побережья Кавказа. Из них можно указать на катилларию Боутелли (Catilaria bouteillei Mass. em. Th. Fr.) и бацидию грушевидную (Bacidia apiahica Koerb.), растущие на верхней поверхности старых листьев чайного куста, что, без сомнения, отражается на общем развитии куста. Катиллария Боутелли встречается также на хвое и живых веточках елей и пихт.
Эпифилльными, хотя и с известной приблизительностью, можно назвать некоторые распространенные листоватые лишайники из родов фисция (Physcia(Schreb.) Vain) и пармелия (РаrmeliaAch.), иногда нарастающие с коры веток на хвою хвойных деревьев.
Эпифитные лишайники на коре деревьев очень многочисленны. Здесь обитают и накипные, и листоватые, и кустистые формы. Нередко они сплошь покрывают ствол дерева на большом протяжении. На участке коры величиной не более ладони иногда насчитывали до 38 видов лишайников, которые росли вплотную друг около друга и даже один на другом.
Существует ли какая-нибудь зависимость флоры лишайников от древесной породы, еще точно не установлено. Правда, наблюдения показывают, что на отдельных породах деревьев часто наблюдаются определенные группировки лишайников. Так, в Подмосковье на осинах чаще встречаются стенная золотнянка (XanthoriaparietinaBelt.), фисция припудренная (Physciapulverulenta(Schreb.) Hampe.), фисция щетинистая (Ph. hispida(Schreb.) Fr.), фисция звездчатая (Рh. stellaris(Ach.) Nyl.), фисция темная, или реснитчатая (Рh. ciliate(Hoffm.) DR.), пармелия бороздчатая (РаrmeliasulcataTayl.); на сосне тАУ уснея длиннейшая (Usnealongissima Ach.), алектория оливковая (Alectoriaolivacea Ach.), рамалина волосовидная (Ramalinathrausta(Ach.) Nyl.); на рябине и дубе тАУ пармелия бороздчатая. Эти же виды могут встречаться и на других породах, хотя и не столь часто.
Для преобладающего расселения того или иного вида лишайников имеет значение строение коры (ее физические свойства, химический состав, кислотность и т. д.). Приуроченность лишайников к определенным древесным породам в известной степени зависит и от климатических условий, в которых произрастает данная порода. Например, различаются по качественному и количественному составу лишайников сосны Прибалтики, средней полосы Европейской части СССР и Европейского Севера.
Распространено мнение, что эпифитные лишайники поселяются на старых, ослабленных деревьях. Действительно, в ряде случаев это так: на старых экземплярах елей более богатый видовой состав лишайников; старые, ослабленные плохим уходом и нарушением агротехники кусты чая также быстрее заселяются эпифилльными лишайниками. Однако лишайники часто растут и на молодых, хорошо развитых деревьях и кустарниках [4].
Расселение лишайников на стволе зависит в основном от освещенности. Лишайники, приспособленные к существованию при малой освещенности, поселяются ближе к основанию ствола, а светолюбивые поднимаются по стволу. На основании ствола лишайники конкурируют с мхами. Вероятно, здесь они частично паразитируют на мхах [10].
Флора эпиксильных лишайников, поселяющихся на обнаженной или обработанной древесине, обычно близка по видовому составу к флоре лишайников, обитающих па коре деревьев, почве и отчасти на скалах.
Эпилитные лишайники поселяются; на камнях и скалах и представлены в основном накипными видами. Расселение их по субстрату различно. Один вид может сплошь покрывать скалу или крупный камень на большой площади, придавая им заметную издали, характерную для определенного вида лишайника желтую, оранжевую, зеленоватую, коричневатую, черную или другую окраску. В другом случае на небольшом участке могут произрастать несколько видов лишайников, образуя на субстрате благодаря своей разнообразной окраске пестрый узорчатый рисунок.
Среди эпилитных лишайников различаются кальцефильные виды, живущие на известковых породах [виды рода веррукария (Verrucaria Wigg. em. Th. Fr.), коллема черная (Collemamultifidum (Huds.) DC.), гаcпарриния обманчивая (Gaspariniadecipiens(Arn.) Sydow.), некоторые виды рода биатора (Вiotora Ach. em. Koerb.) и др.], и кал
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)