Отдел голосеменные
Отдел диатомовые водоросли
Сюда относятся коккоидные тАФ одноклеточные и колониальные водоросли с хроматофорами, в живом состоянии окрашенными в желто-бурый цвет благодаря преобладанию каротиноидов над хлорофиллами а, с. Хлоропласта обычно париетальные, в виде одной-двух крупных пластинок или многочисленных дисков. Мат-рикс хлоропласта пересекается трехтилакоидными ламеллами, имеется опоясывающая ламелла. Пиреноид центральный, в большинстве случаев окружен специальной мембраной и пронизан различным у разных видов числом тилакоидов. Запасные продукты тАФ хризоламинарин, масло, волютин; крахмал отсутствует. Клеточное ядро локализовано или в постенной цитоплазме, окружающей одну центральную вакуоль с клеточным соком, или в цитоплазматичес-ком мостике, пересекающем вакуоль. Цитоплазма содержит пери-нуклеарный комплекс диктиосом, митохондрии. Монадные клетки одножгутиковые, в аксонеме отсутствуют центральные микротрубочки.
Клеточный покров. Диатомовые характеризуются особым клеточным покровом, состоящим из двух главных компонентов: кремнеземного панциря и тонкого органического пектинового слоя, который полностью обрамляет панцирь. Для изучения панциря диатомовые обрабатывают таким образом, чтобы разрушить протопласт, при этом удаляют и поверхностный органический материал. Чтобы изолировать второй компонент клеточного покрова диатомовых тАФ органический наружный слой, на клетку воздействуют фтористоводородной кислотой, растворяющей кремнезем. Топография такого органического покрова оказалась видоспецифичной и идентичной с топографией панциря. Пока неясно, отражает ли рисунок органического покрова рисунок панциря или же, наоборот, является матрицей для панциря. Органический слой при формировании клеточного покрова развивается интрацеллюлярно, у одних диатомей он может утрачиваться, у другихтАФ сохраняется. Кроме того, через отверстия в панцире может выделяться слизь, которая в зависимости от вида формирует слизистые чехлы, трубки, ножки, подушечки, тяжи, сосочки и пр.
Панцирь диатомовых водорослей состоит из двух половинок: большей эпитеки и меньшей гипотеки. Каждая половинка образована створкой, которая своим загибом соединяется с поисковым кольцом или ободком. Свободный край пояскового кольца эпитеки надвинут на свободный край пояскового кольца гипотеки, как крышечка на коробочку. У многих диатомей между загибом створки и поясковым кольцом возникают вставочные ободки различной формы и в разном количестве. Эти ободки появляются последовательно, причем каждый вставочный ободок формируется между загибом створки и последним из ранее образовавшихся вставочных ободков. Таким образом самый молодой вставочный ободок всегда граничит с загибом створки, а самый старый тАФ с поясковым ободком. На вставочных ободках развиваются септы тАФ неполные кремнеземные перепонки, расположенные или вдоль всей внутренней поверхности вставочного ободка, или на одном из его полюсов. Септы всегда имеют одно или несколько отверстий, перегораживая полость клетки на сообщающиеся между собой отсеки. Образование вставочных ободков обусловливает возможность роста клетки вдоль ее центральной оси, соединяющей центры двух створок. Различные части панциря плотно сцементированы вместе, иногда связаны сложными закраинами. Раздвигание эпи- и гипотеки наблюдается только при делении клетки и во время образования ауксоспор. Створки у одних диатомовых, объединяемых в класс центрофицие-вые, обычно округлых очертаний, например у рода циклотелла, и панцирь в целом может быть уподоблен круглой коробочке. Реже створки треугольные, как у трицера-циума, или эллиптические, как у рода биддульфия. Элементы скульптуры, представленные в первую очередь порами различных типов, размещаются на створках вокруг центральной точки, а у родов с округлыми створками обычно располагаются радиально. У других диатомовых, составляющих класс пеннатофициевых, створки эллиптические или продолговатые, например у родов пин-нулярия и навикула, у некоторых, как у синед-ры, длина их во много раз превосходит ширину.
У перечисленных родов концы створок одинаковые тАФ изополъ-ные створки. У родов гомфонема, ликмофора

створки гетерополъные, т. е. суживаются к одному концу. Могут быть створки и иной формы: S-образно изогнутымитАФ у плевроеигмы, гиросигмы {Gyrosigma), некоторых видов рода ницшия, полулунными с прямым или вогнутым брюшным и выпуклым спинным краями, как у цимбеллы iCymbdla) и т. д. У всех пеннатофициевых скульптурные элементы на створках располагаются в два продольных ряда по бокам от бесструктурной срединнойтАФмедианной линии, или осзвого поля. Положение клетки диатомовых, при котором видна одна из створок, называется видом панциря <_о створки. Кроме того, различают вид панциря с пояска, при котором видно, как поисковое кольцо эпитеки надвинуто на поисковое кольцо гипотеки. У громадного большинства диатомовых, цен грофициевых и пеннатофициевых клетка со стороны пояска имеет форму четырехугольника, но поясок может быть и клиновидным, S-образно изогнутым с более или менее выраженным изгибом посередине тАФу рода ахнантес {Achnanthes) и пр.
У некоторых пеннатофициевых, объединяемых в порядок фраги-ляриальные, осевое поле тАФ бесструктурная часть створки тАФ гладкое. Этим характеризуются роды синедра, меридион {Meridian), астерионелла, фрагилярия, диа-тома. У большинства других пеннатофициевых эта область створки занята особым образованием тАФ швом_, имеющим вид двух продольных, слегка изогнутых щелей в толще створки. Это щелевидный шов. В середине створки, где панцирь изнутри утолщен, образуя центральный узелок, обе ветви шва погружаются в две центральные поры в толще центрального узелка. У полюсов клетки каждая ветвь шва заканчивается внутри полярныхтАФ конечных узелков. Последние, как и центральный узелок, являются местными, но менее заметными утолщениями створки На поперечном разрезе створок можно видеть, что шов представляет собой V-образную или коленовидно изогнутую трещину, состоящую как бы из двух щелей или каналов, из которых одна открывается в полость клетки, а другая тАФ наружная тАФ сообщается с внешней средой. Наружная и внутренняя щели каждой ветви шва соединяются в центральном узелке посредством вертикального канала. В центральном же узелке обе ветви шва сообщаются с помощью горизонтального канала.
Внутри полярных узелков наружная щель шва переходи! в слегка скрученную трещину, а внутренняя щель заканчивается в виде воронковидиого образования.
Описанный выше щелевидный шов развивается на обеих или только на одной створке пеннатофициевых, объединяемых в порядок навикулальные. Каналовидный шов характерен для диатомовых, относящихся к порядку суриреллальные. Он не связан с осевым полем створки, обычно располагается в более или менее выступающем гребне или киле, идущем продольно вдоль середины створки или лежащем эксцентрично ближе к одному из ее краев. Каналовидный шов представлен обычно непрерывным трубковидным каналом в толще створки, сообщающимся с наружной средой посредством тонкой щели, гомологичной наружной щели щелевидного шва. С внутренней полостью клетки канал соединяется через отверстия или поры, гомологичные внутренней щели щелевидного шва. Диатомовые, имеющие шов, способны к движению. Механизм движения окончательно не выяснен. Имеется ряд гипотез. По одной, наиболее распространенной гипотезе движение клетки тАФ следствие трения о субстрат цитоплазмы, циркулирующей в шве. Благодаря внутриклеточному тургору цитоплазма вгоняется в вертикальный канал центрального узелка, откуда по внутренней щели шва гонится к полярным узелкам, а там по воронкообразному каналу передвигается в конечную щель полярного узелка и попадает в наружную щель шва, по которой течет по направлению к центральному узелку. В центральном узелке по вертикальному каналу, соединяющему наружную и внутреннюю щели, этот ток

цитоплазмы может перейти во внутреннюю щель этой же ветви шва, чтобы повторить пройденный путь, но может попасть и непосредственно в полость клетки или же через горизонтальную борозду, соединяющую в центральном узелке обе ветви шва, переходит во внутреннюю щель другой ветви шва и начинает там круговой путь в противоположном направлении. Трение струящейся цитоплазмы о субстрат и развивает моторную силу, движущую клетку в направлении, противоположном току цитоплазмы. Движение форм, имеющих каналовидный шов без центрального узелка, значительно интенсивнее благодаря непрерывному току цитоплазмы.
Согласно другой гипотезе скольжение диатомовых обусловлено движением жидкости в шве, представляющем собой пластинчатый капилляр, в который жидкость втягивается под действием капиллярных сил.
У имеющих шов пеннатофициевых хлоропласты располагаются на поисковой стороне клетки, лишь слегка заходя на створковую. Перед делением клетки хлоропласты перемещаются на створку. У бесшовных форм хлоропласты располагаются со стороны створки.
Ничего сравнимого со швом не наблюдается у представителей центрофициевых. Они все неподвижны.
Многие диатомей в течение всей жизни встречаются в виде отдельных клеток, однако известны и колониальные формы, как свободноплавающие, так и прикрепленные. Колонии берут свое начало от одной клетки, которая в результате ряда делений дает свойственную данному виду форму колонии. Как правило, колонии образуются у видов, не способных к движению, т. е. совсем не имеющих шва. Виды же с вполне развитым щелевидным или каналовид-ным швом формируют колонии редко, и при этом клетки, входящие в состав колонии, обычно не теряют способности к движению.
Клетки прикрепляются к субстрату и соединяются в колонии с помощью слизи, выделяемой через сравнительно крупные поры на поверхности створки. Иногда в скреплении клеток в колонии, помимо слизи, принимают участие различные выросты створоктАФрога, щетинки, зубчики и пр. Все громадное разнообразие форм колоний можно свести к трем основным типам. К первому типу относятся студенистые колонии, в которых клетки целиком погружены в выделяемую ими слизь. Колонии имеют вид бесформенных комочков или трубок, неэетвящихся или разнообразно разветвленных.
Второй тип включает нитевидные, лентовидные, цепочковидные и звездчатые колонии, форма их зависит от способа скрепления клеток в колонии и от формы створок. Во всех случаях клетки соединяются только створками, соединения поясками никогда не бывает, так как в плоскости пояска клетки делятся. Если смежные клетки соединяются всей поверхностью створки, то в зависимости от формы створок образуются колонии нитевидные, как у некоторых видов циклотеллы и у рода мелозира, лентовидныетАФ например у фрагилярии, вееровидные тАФ у меридион. Если же клетки смыкаются только концами створки, то возникают или зигзагообразные цепочки, как у рода диатома, или звездчатые колонии, характерные для рода астерионелла
У различных планктонных центрофициевых клетки соединяются выростами створок, слизистыми тяжами и тогда образуются не плотные, а рыхлые цепочки, в которых клетки более или менее удалены друг от друга. Таковы роды хетоцерос, скелетонема {Skeletonema, рис. 27, АтАФВ), талассиози-ра.
Третий тип колоний наблюдается у форм, прикрепленных к субстрату слизью, которая выделяется локально на базальном конце клетки через поры. Слизь имеет вид тяжей неразветвленных или древовидно ветвящихся, на концах которых сидят клетки тАФ по одной или помногу. Примерами могут служить виды синедры, гом-фонемы, ликмофоры.
Электронно-микроскопические исследования панциря диатомовых водорослей показали, что кремневые стенки панциря бывают однослойными и двухслойными независимо от принадлежности данного вида к центрофициевым или пеннатофициевым диатомовым. В первом случае стенка панциря состоит из одной пластины, во второмтАФиз двух пластин: наружной и внутренней, пространство между которыми разделено вертикальными перегородками на камеры. Кроме того, было показано, что створки панциря всегда пористые, и один из основных элементов структуры панциря тАФ именно поры. По классификации X. Окуно различают три основных типа пор: отверстия, локулы, альвеолы. Отверстия тАФ это мелкие, в виде следа булавочного укола округлые или щелевидные перфорации, возникающие в более или менее тонких однослойных стенках. Отверстия лишены замыкающих ситовидных мембран. В двухслойных стенках встречаются или округлые и полигональные перфорации локулы, или более или менее вытянутые перфорациитАФ альвеолы. И те и другие снабжены замыкающими ситовид-

иыми мембранами, перфорированными, в свою очередь, ситовидными порами. Локулы могут иметь хорошо выраженные высокие латеральные стенки, которые вдаются глубоко внутрь, и тогда ситовидная мембрана находится снаружи локулы; если латеральные стенки высоко выступают наружу, то ситовидная мембрана расположена у основания локулы. В первом случае изнутри, а во втором--снаружи локул обычно наблюдается покровная мембрана, перфорированная всегда одной крупной поройтАФ покровной порой. В том и другом типе локул покровная мембрана может отсутствовать. Латеральные стенки бывают с латеральными порами или без них. В толстых стенках, перфорированных удлиненными альвеолами, как и в случае локул, латеральные стенки вдаются внутрь или выступают кнаружи, соответственно ситовидная мембрана располагается снаружи от альвеолы или у ее основания. Покровные мембраны могут присутствовать или отсутствовать. Пор в латеральных стенках альвеол до сих пор не наблюдалось.
Ситовидные мембраны, которые замыкают локулы или альвеолы на их наружной или внутренней поверхностях, имеют характерную тонкую структуру. Ситовидная мембрана перфорирована одной или многими крошечными ситовидными порами округлых, эллиптических, прямоугольных или полигональных очертаний. Ситовидные поры, в свою очередь, часто замкнуты тонкой вторичной мембраной с ультратонкими вторичными ситовидными порами. Форма, размер и расположение ситовидных пор могут служить в качестве надежного признака для идентификации видов.
Несколько подробнее можно рассмотреть структуру панциря у рода пиннулярия. В панцире пиннулярии альвеолы поперечно вытянуты, простираясь почти от шва до края створки. Они представлены корытообразной полостью, замкнутой снаружи только пористой ситовидной мембраной. У некоторых видов пиннулярии альвеолы, лишенные покровной мембраны, полностью открыты внутрь, у других видов альвеолы изнутри загорожены тонкой покровной мембраной с отверстиемтАФ покровной порой. Покровные поры могут быть округлыми, эллиптическими или другой формы, они расположены на створке в две продольные линии. Эти ряды покровных пор различимы и в световом микроскопе. Поперечные боковые стенки альвеол более или менее глубоко вдаются внутрь и лишены пор. Таким образом, соседние альвеолы полностью разделены этими боковыми стенками.
Электронно-микроскопическая тонкая структура пор панцирей диатомовых водорослей строго видоспецифична. Согласно X. Оку-но, Влнадежность тонкой структуры пор панциря для идентификации видов диатомовых водорослей может быть сравнима с надежностью отпечатков пальцев для идентификации человеческих индивидуумовВ».
Значение пор, по-видимому, заключается в том, что через них содержимое клетки сообщается с окружающей средой.
Во многих руководствах и исследованиях можно встретиться и с несколько иной терминологией. Различают два основных элемента структуры панциря: поры и ареолы. В однослойных тАФ ламинарных стенках образуются только поры, которые могут быть открытыми в виде сквозного отверстия или полузамкнутыми мембранами разного типа. Ареолы образуются только в двухслойных тАФ локу-лярных стенках и обычно затянуты снаружи или внутри тонкой перфорированной пленкойтАФВлвелумомВ» или ВлкрибрумомВ». Помимо пор, как уже указывалось, на створках имеются различною рода выросты, характер и расположение которых служат важными таксономическими признаками. Так у ряда центрофи-циевых диатомей существуют выросты: центральные, краевые, промежуточные между ними и одиночный или добавочный вырост, расположенный или между маргинальными выростами, или же между центром и краем створки. Г. Хасле подробно изучила их строение и назначение. Центральные выросты оказались полыми трубками, иногда расширенными на вершине, основание их окружено ободком в виде муфты, прерванной в четырех местах, что позволяет видеть стенки цилиндров до самого основания. Хасле назвала эти трубки Влподпертыми трубкамиВ». Их функция состоит в выделении плотной слизи, образующей студенистые тяжи, соединяющие смежные клетки в це-почковидную колонию. Краевые и промежуточные выросты также трубчатые, открытые наружу и внутрь клетки, через них выделяется слизь, покрывающая всю поверхность створки. Эти выросты свойственны только планктонным видам. Крупный одиночный вырост тоже трубчатый, иногда на вершине он расширен, а внутрь клетки открывается узкой щелью с утолщенными краями в виде двух губ. Хасле назвала эти выросты двугубыми; функция их пока не ясна. Примерами строения и назначения этих выростов могут служить выросты у представителей родов талассиозира и ске-летонема {Skeletonema).
У большинства видов талассиозиры клетки образуют рыхлые цепочки посредством слизистого тяжа, соединяющего центры смежных створок. С помощью сканирующего электронного микроскопа показано, что эта соединительная нить между соседними панцирями образована несколькими нитями, которые выдавливаются из 7тАФ12 Влподпертых трубокВ», расположенных в центре створки. Помимо центральных подпертых трубок, на створке имеются более мелкие подпертые трубки и один двугубый отросток. У разных видов они расположены по-разному. Отличительный признак рода скелетонема {Skeletonema) тАФ соединительные отросткитАФ маргинальные подпертые трубки в числе 7тАФ9 со срав-
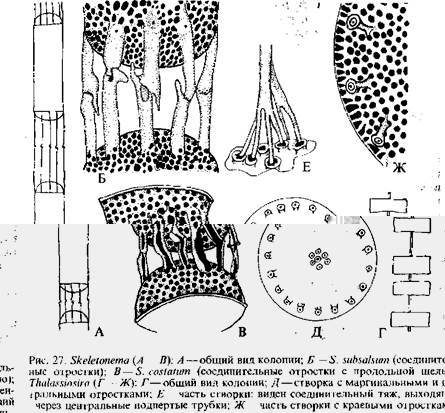
нигельно длинными наружными частями, которые смыкаются обычно по одному с такими же отростками смежных клеток, образуя рыхлые цепочки. Структура соединительных отростков у разных видов неодинакова. Так, у S. subsalsum они трубчатые, а у 5*. costatum расщеплены в виде желоба почти на протяжении всей длины.
Помимо трубковидных, створки могут нести и иные выросты, часто также служащие для объединения клеток в колонии. Например, у рода хетоцерос с цепочкообразными колониями клетки соединяются с помощью длинных негнущихся щетинок. Овальные створки биддульфии тоже снабжены выростами, которые объединяют клетки в рыхлые цепочки.
Размножение. Обычный способ размножения у диатомей тАФ клеточное деление. В результате этого процесса оба образующихся индивида получают одну створку Т>т родительской клетки, тогда как другая возникает заново. Две створки в делящейся клетке слегка расходятся так, что остаются соединенными только края их поисковых ободков. Ядро претерпевает митоз. Расщепление протопласта начинается с периферии, идет ковнутри и всегда происходит в плоскости, параллельной створкам. На заново образовавшейся свободной поверхности обоих дочерних протопластов формируются новые створки, а затем около каждой из новых створок возникает поисковый ободок и всегда ковнутри от поисковых ободков первоначального индивида. Таким образом, поисковый ободок каждой родительской створки перекрывает поисковый ободок заново образовавшейся створки, и гипотека родительской клетки становится эпитекой одного из дочерних индивидов. Возникновение новых створок можно проследить только с помощью электронного микроскопа. Электронно-микроскопические исследования показали, что в каждой полученной в результате деления клетке, окруженной плазмалеммой, недостающаи створка формируется внутри периферического плоского, окруженного мембраной пузыря тАФ Влпузыря отложения кремнеземаВ», который служит матрицей, определяя окончательную морфологию створки. Пузырь отложении кремнезема возникает под плазмалеммой, по-видимому, благодаря слиянию пузырьков Гольджи. Мембрана, ограничивающая этот пузырь, получила название силиколеммы; возможно, она ответственна за отложение кремнезема, т. е. за транспорт иона SiO^"2 в пузырь и его полимеризацию там до кремнезема. По достижении новой створкой зрелости и ее освобождении силиколемма, обращенная внутрь, становится плазмалеммой' и, возможно, составляет часть органического покрова, возникающего в основном за счет слияния силиколеммы и плазмалеммы, обращенных кнаружи клетки.
Ниже приводится несколько упрощенная схема процесса образования новых половинок панцири в делящейся клетке Gomphonema parvulutn. Сравнительно недавно у Fragiiaria capucina было прослежено возникновение иитеркалярных тАФ вставочных поясков. Было показано, что они также формируются внутри пузырей отложения кремнезема, возникающих непосредственно под плазмалеммой, по-видимому, в результате слияния везикул- дериватов аппарата Гольджи. Ободки покрыты матриксом, который удерживает их вместе. Таким образом, элементы панцири являются независимыми единицами, которые и формируются в сепаратных Влпузырях отложения кремнеземаВ». Этим подтверждаются более ранние данные световой микроскопии: поясковые и вставочные ободки не образуются до тех пор, пока не будет завершено формирование створки.

В результате деления клетки одна из дочерних клеток, получившая от материнской эпитеку, точно повторяет размеры материнской клетки. Вторая дочерняя клетка, получившая от материнской гипотеку, которая стала для нее эпитекой, несколько меньших размеров: на двойную толщину пояскового ободка. Из-за окре-мнения панциря клетки могут увеличиваться только вдоль оси, соединяющей две створки. В результате размеры клеток в популяции диатомовых прогрессивно уменьшаются. Скорость этого уменьшения размеров не так уж велика, некоторое растяжение панциря у слабо окремнелых диатомей все же наблюдается и частота делений, особенно меньших из двух дочерних клеток, невелика. Специальное исследование показало, что уменьшение размеров клеток до минимальных может растягиваться на несколько лет: на два тАФ пять у Stephanodiscus hantzschii, на четыре года у Tabellaria fenestrata. Затем следует краткий период образования растущих спор тАФ ауксоспор, возникающих в результате полового процесса. Зигота, одетая неокремнелой растяжимой оболочкой, превращается в ауксоспору, которая растет, пока не достигнет максимальной для данного вида величины, после чего образуется панцирь и начинается ряд последовательных вегетативных делений клеток.
У пеннатофициевых половой процесс напоминает конъюгацию, когда сливаются протопласты вегетативных клеток. В качестве примера можно рассмотреть половой процесс у Gomphonema geminatum, подробно изученный русским аль-гологом К. И. Мейером. Две клетки сближаются и окружаются толстой слизистой обверткой. Ядро каждой из конъюгирующих клеток претерпевает редукционное деление, а протопласт делится с образованием двух гамет одинакового размера. Гаметы сначала содержат по два ядра, одно из которых остается жизнеспособным, а другое дегенерирует. После этого створки конъюгирующих клеток расходятся, и одна гамета из каждой клетки переползает в клетку партнера с тем, чтобы слиться там с другой гаметой. Возникшие в результате конъюгации две зиготы начинают быстро расти и превращаются в ауксоспоры. Ауксоспоры сначала двухъядерные, но вскоре происходит кариогамия и более зрелые ауксоспоры содержат по одному диплоидному копуляционному ядру. Разрастаясь, они растягивают, а затем и разрывают общую слизистую обвертку, одевающую конъюгантов, в которой можно видеть створки конъюгирующих клеток, значительно меньших размеров, чем ауксоспоры. Достигшая максимальных размеров ауксоспора образует панцирь со створками, несколько различающимися между собой: появляющаяся первой тАФ более толстая и выпуклая, другая створкатАФплоская. Только после деления ауксоспоры дочерние клетки приобретают признаки, свойственные вегетативным клеткам данного вида. У большинства пеннатофициевых половой процесс протекает таким же образом, т. е. в каждом из конъюгантов образуется по две гаметы и в результате возникает по две ауксоспоры. В деталях могут быть и различия. Например, у Anamoeoneis sculpta деления в конъюгантах неравновеликие, образуются по две гаметы, различающиеся по размерам. Затем гаметы разного размера из разных конъюгантов попарно сливаются. Реже у пеннатофициевых редукционное деление ядра в конъюгирующих клетках не сопровождается цитокинезом и таким образом образуется по одной гамете, содержащей сначала четыре ядра, из которых три затем отмирают. В результате такого полового процесса формируется только одна зигота, соответственно тАФ ауксоспора. У некоторых пеннатофициевых известны случаи апогамии, когда ауксоспоры возникают из одной-единственной родительской клетки. Здесь сливаются два ядра из четырех получившихся в процессе редукционного деления в неразделившихся протопластах, которые превращаются в зиготы, вырастающие в ауксоспоры.
Таким образом, у пеннатофициевых имеет место гаметическая редукция: перед образованием гамет вся вегетативная жизнь их проходит в диплоидном состоянии тАФ они являются диплонтами.
В начале XX в. проблема изучения жизненного цикла центрофициевых усложнилась благодаря обнаружению сначала у рода биддульфия, а позднее и у других родов монадных клеток тАФ микроспор. Ряд авторов приписывали им роль гамет и соответственно отрицали половой акт при образовании ауксоспор. В 1929 г. русский исследователь Б. М. Персидский, соглашаясь с тем, что ауксоспоры центрофициевых возникают из одной клеткитАФматеринской клетки ауксоспор, без слияния ее с другой клеткой, отрицает бесполый характер ауксоспор. У двух видов рода хетоцерос и у Melosira varians Б. М. Персидский наблюдал в материнской клетке ауксоспор редукционное деление ядра и образование четырех гаплоидных ядер. Он утверждал, что два из четырех ядер отмирают, а два оставшихся сливаются в молодой ауксоспоре, которая возникает таким образом в результате апогамного полового процесса. Позднее то же наблюдалось у Cyclotella meneghiniana. На основе этих данных до 1950 г. господствовало мнение, что у центрофициевых ауксоспоры образуются из одной клетки, в которой происходит редукционное деление ядра, и два ядра сливаются в созревающей ауксоспоре; этот процесс сравнивался с встречающейся у некоторых пеннатофициевых аутогамией. Однако в 1950тАФ1954 и последующие годы появляются работы, в которых у различных представителей центрофициевых описывается оогам-ный половой процесс. Первая работа Г. Штош относилась к пресноводной водоросли Melosira varians тАФ виду, изученному Б. М. Персидским. Штош показал, что ауксоспора М. varians является разросшейся зиготой, получающейся в результате слияния сперматозоида с материнской клеткой ауксоспоры. Согласно Штош, антеридии возникают в результате двух, реже одного, трех или четырех делений из вегетативных клеток. В ан-теридиях происходит редукционное деление, вокруг получившихся четырех гаплоидных ядер обособляется цитоплазма и образуются одножгутиковые, лишенные хроматофоров сперматозоиды. Антеридии и оогонии могут встречаться на протяжении одной и той же нити. По мнению Штоша, Б. М. Персидский ошибочно отождествил антеридии с четырьмя ядрами как стадию тетрады, предшествующую автогамной кариогамии при образовании ауксоспор у мелозиры.
Обнаружение подвижных монадных сперматозоидов у мелозиры позволило предположить, что хорошо известные микроспоры многих морских центрофициевых также являются сперматозоидами и что и у них образованию ауксоспор предшествует оогамия. Это предположение подтвердилось и в последующие годы. Штош устанавливает оогамию у целого ряда морских центрофициевых: Lithodesmium, Bellarochea, Streptotheca, Biddulphia rhombus, В. granulata, В. mobiiiensis, Cerataurus, Aulacodiscus. Л. Гей-тлер описал оогамию у циклотеллы.
В деталях половой процесс и ово- и сперматогенез у разных видов неодинаков, В материнской клетке ауксоспор тАФ оогонии после редукционного деления ядра могуч образоваться два яйца или же в оогонии формируется только одна яйцеклетка. Первый путь овогенеза характерен, например, для В. mobiiiensis, В. granulata, Lithodesmium. Второй тип наблюдается у Melosira varians, Cydotella tenuistriata, Streptotheca. Промежуточное положение занимает В. rhombus, у которой в оогонии имеет место неравное деление протопласта, в результате которого образуется крупная яйцеклетка и мелкое, лишенное хлоропластов так называемое направляющее тельце. В первом случае в результате полового процесса формируются две ауксоспоры, во втором тАФ только одна. Яйцеклетки, возникающие в оогонии в числе двух или одной, в одних случаях оплодотворяются внутри оогония, т. е. происходит истинная оогамия, в других тАФ только после выхода яйцеклеток из оогония, т. е. наблюдается примитивная оогамия. В антеридиях все четыре ядра, возникающие в результате редукционного деления исходного диплоидного ядра, остаются жизнеспособными и вокруг них обособляются четыре сперматозоида. Однако дифференцировка самих антеридиальных клеток у разных представителей происходит неодинаково. У Cydotella tenuistriata вегетативные клетки непосредственно становятся антеридиями. У Melosira varians из одной вегетативной клетки возникают 4тАФ8 более мелких, с несколько упрощенными панцирями, бедных хлорофиллом сперматогониев. У Biddulphia rhombus и В. granulata сперматогонии, образующиеся из вегетативной клетки, более редуцированы: одеты сильно упрощенными панцирями и до мейоза заключены в материнской клетке. У В. mobiliensis после ряда митозов из вегетативной клетки появляются 16 ан-теридиев, которые освобождаются после расхождения створок, образующихся только после первого митоза. То же имеет место и у лито-десмиума, стрептотеки и некоторых других, у которых отсутствуют все створки, кроме сформировавшихся после первого митоза, так что голые сперматогонии, возникающие в больших количествах, заканчивают свое развитие свободно в окружающей среде. v
Штош наблюдал слияние сперматозоида с яйцом у В. mobiiiensis на живом материале, а также представил цитологические доказательства слияния ядер. Он также продемонстрировал экспериментально, что яйца способны прорастать в ауксоспоры только в присутствии сперматозоидов. В 70-х годах подробно описан жизненный цикл и оогамный половой процесс у нескольких видов хетоцерос. Оогамия описана лишь у одного вида пеннатофициевых диатомовых тАФ Rhahdonema adriaticum. Однако мужские клетки здесь лишены жгутика и они передвигаются посредством тонких псевдоподий.
Вегетативные клетки большинства диатомовых водорослей способны выносить высыхание, низкие температуры и прочие неблагоприятные условия. Однако у некоторых форм, преимущественно морских центрофициевых, реже у пресноводных видов, известны покоящиеся споры или цисты. Образованию их обычно предшествует деление клетки, при этом дочерние клетки покрываются толстостенными выпуклыми створками, смыкающимися своими краями, поясковые кольца отсутствуют. Створка, возникающая первой, покрыта шипами или выростами, створка, "образующаяся позже, гладкая.
* * *
Критерии, которые кладутся в основу систематического деления диатомовых водорослей на классы, порядки, семейства и т. д., касаются формы створок, деталей их структуры, отсутствия или наличия вставочных ободков, шва, их формы и т. п. Как уже упоминалось, отдел диатомовые водоросли делится на два класса: центро-фициевые и пеннатофициевые.
КЛАСiЕНТРОФИЦИЕВЫЕ тАФ СENTROPНYСЕА Е
Створки обычно округлых очертаний и радиального строения, шов отсутствует, половой процесс тАФ оогамия. Примерами могут служить роды циклотелла, мелозира, талассиозира, скелетонема, биддульфия, хетоцерос.
КЛАСС ПЕННАТОФИЦИЕВЫЕ тАФPENNATOPHYCEAE
Створки вытянутые, с двусторонней симметрией. У одних шов щелевидный, что характерно для порядка навикулальные, или каналовидный, у других шов отсутствует тАФ порядок фрагиляриальньге. Половой процесс по типу конъюгации. К пеннатофициевым относятся роды пиннулярия. на-викула, цимбелла, гиросигма, гомфонема, ахнантес; ницшия, сурирелла; синедра, фрагилярия, диатома, астерионелла, меридион.
Оба классатАФцентрофициевые и пеннатофициевыс, по-видимому, произошли от одного общего предка, в пользу чего говорит диплоидная природа обеих групп, сходство строения панциря, пигментации и обмена всшсств. Центрофициевые и лишенные шва пеннатофициевыс могут представлять собой дивергирующие линии эволюции от этого общею ствола, в то время как остальные пеннатофициевые, главным образом бентосные формы, эволюционировали в сторону выработки сложного строения шва. Из двух классов центрофициевые больше сохранили черт, связывающих их с какими-то отдаленными жгутиковыми предками, в пользу чего свидетельствует наличие у них жгутиковых стадий.
* * *
Диатомовые встречаются во всех видах вод, а также во многих типах почв. Мно1ие несущие шов виды являются донными формами, живущими в пресной и соленой воде, другие -эпифиты тАФ на более крупных растениях. Диатомовые,.чишенные шва, играют большую роль в планктоне, причем в море преобладают центрофициевые, а в пресноводном планктоне фрагиляриальные. В качестве создателей органического вещества в водоемах диатомовые занимают среди всех водорослей первое место. Поскольку сильно окремневшие оболочки сохраняются неизменными после отмирания водорослей, то в местах массового развития диатомей образуются обширные отложения, известные как диатомовая земля или кизельгур, имеющие важное экономическое значение: как полировочный материал, для тепловой изоляции, как связующая масса при изготовлении динамита. В меловых отложениях диатомовые представлены большим количеством видов. Совершенство строения меловых диатомей заставляет предполагать, что они уже до мелового периода прошли длительную эволюцию.
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)