Постсинаптическая трансформация сигнала
Активность нервных окончаний регулируется двумя экзогенными факторами тАУ изменением мембранного потенциала и прямым взаимодействием медиаторов нервных импульсов с рецепторами. В результате этих событий меняется цитоплазматический уровень по меньшей мере вторичных посредников тАУ Са+, цАМФ, цГМФ, инозитолтрифосфата и диацилглицерина, что приводит к активации соответствующих пулов протеинкиназ: цАМФ-зависимых протеинкиназ; цГМФ-зависимых протеинкиназ; Са-кальмодулин и Са-фосфолипид-зависимых протеинкиназ. Активация протеинкиназ обусловливает фосфорилирование регуляторных белков-мишеней в клетках нервной системы и тем самым модулирует функциональную активность этих клеток.
Определяющий вклад в проблему внутриклеточной регуляции был сделан в 50тАУ60-е годы Э. Сазерлендом, сформулировавшим представление о роли циклических нуклеотидов как вторичных посредников, накапливающихся в клетке в ответ на нейромедиаторный или гормональный стимул и осуществляющих связь между рецепторами и исполнительными системами. В результате дальнейшего развития этих исследований оказалось, что циклический аденозинмонофосфат регулирует обмен белков, углеводов, липидов и нуклеиновых кислот, влияет на проницаемость мембран, электрическую, сократительную и секреторную функции клеток, дифференцировку и пролиферацию. Установлена роль фосфорилирования белков как основного пути действия этого нуклеотила на клетки животных. Найдена связь между содержанием циклических нуклеотидов и характером протекания некоторых патологических процессов в тканях. Описано участие цАМФ и цГМФ в проявлении действия многих лекарственных препаратов на организм. Спустя лишь 10тАУ15 лет после открытия, сделанного Э. Сазерлендом, представления о роли и механизме действия циклических нуклеотидов оказались неотделимыми от современной биохимии, физиологии и медицины. Задача настоящей работы состоит в сравнительно детальном рассмотрении постсинаптических механизмов.
1. цАМФ-Зависимое фосфорилирование
1.1 Аденилатциклаза, ее активация
Синтез в клетке цАМФ из АТФ осуществляет аденилатциклаза. Фермент обнаружен практически во всех тканях млекопитающих. Максимальная активность аденилатциклазы выявлена в мозге, далее в порядке убывания активности фермента ткани можно распределить следующим образом: селезенка, скелетные мышцы, сердце, легкие, почка, печень, жировая ткань. Аденилатциклаза локализована почти исключительно в плазматических мембранах, хотя есть сообщения о присутствии фермента в митохондриальной и микросомальной фракциях.
Аденилатциклаза представляет собой мультимолекулярный комплекс, состоящий из рецепторного и каталитического компонентов. Ответ клетки на действие гормона или биологически активного вещества зависит от концентрации рецепторов на поверхности мембраны и степени сопряжения рецепторов с аденилатциклазой. В последнем случае в роли сопрягающего компонента выступают G-белки, обладающие как ГТФ-связывающей, так и ГТФ-гидролизующей активностью.
Рецептия с участием G-белков отличается от других известных систем трансформации внеклеточного сигнала. Все эти системы включают рецептор тАУ дискриминатор сигнала, с высокой специфичностью и чувствительностью "снимающий" внеклеточный сигнал с наружной мембраны. Рецептор может находиться внутри клетки тАУ в том случае, если эффекторы являются липофильными молекулами, легко проникающими через мембрану. Рецепторы для водорастворимых эффекторов встроены в наружную мембрану: эти рецепторы могут быть ферментами; другой тип мембрановстроенных рецепторов сопряжен с ионными каналами. Наконец, рецепторы, которым не присущи свойства ни канала, ни фермента, сопряжены с ферментами-исполнителями с помощью G-белков.
Регуляцию аденилатциклазной системы с участием G-белков можно представить следующим образом. В системе есть два белка, состоящих из субъединиц а, р и у и названных Gs и Gj. р тАУ у-субъединицы в отличие от а-субъединиц практически идентичны во всех изученных G-белках. а-субъединица содержит участок связывания с гуаниловыми нуклеотидами и обладает ГТФазной активностью. Каждый из G-белков связан со своим рецептором. При образовании комплекса агонистрецептор происходит активация G-белка путем замены в нуклеотидсвязывающем центре ГДФ на ГТФ. При этом изменяется конформация а-субъединицы, что обусловливает ее диссоциацию от комплекса ру. Для диссоциации необходим Mg+. Активированная а-субъединица G| ингибирует активность аденилатциклазы, в то время как активированная Gs тАУ стимулирует ее.
Очевидно, почти идентичные по структуре комплексы Ру могут ингибировать аденилатциклазу непрямым способом, а именно, путем связывания с ct-субъединицей Gs, таким образом "выключая" ее способность стимулировать фермент. Действительно, во многих исследованных тканях, в том числе в нервной, концентрация Gj значительно выше, чем Gs. Следовательно, возможно высвобождение достаточного количества комплекса Ру из Gi для соединения со всеми а-субъединицами Gs. Сопрягающий эффект G-белков прекращается медленным гидролизом связанного ГТФ до ГДФ. G-белки могут быть снова активированы при связывании рецептора с "сигнальной" молекулой.
Установлено, что рецепторы многих гормонов собраны в группы, кластеры и малоподвижны в мембране. Поэтому при активации кластера, состоящего из рецепторов одного типа, происходит стимулирование аденилатциклазы и увеличение уровня цАМФ в ограниченном районе клетки. Это ведет к повышению протеинкиназной активности и фосфорилированию субстратов только в данной зоне клетки, в данном компартменте. Компартментализация цАМФ и А-киназы установлена во многих типах клеток: например, стимуляция сердца как с помощью катехоламинов, так и простагландина Ej приводит к увеличению уровня цАМФ и активации протеинкиназы А. Однако катехоламины и простагландин Ej оказывают различное влияние на фосфорилирование фосфорилазы гликогена. Видимо, рецепторы катехоламинов и простагландина и соответствующие постсинаптические системы расположены в разных компартментах клетки.
1.2 Протеинкиназа
Через 10 лет после открытия Сазерлендом цАМФ была обнаружена цАМФ-зависимая протеинкиназа, значительно повышающая скорость фосфорилирования субстратов в присутствии цАМФ. Реакция протеинфосфорилирования выглядит следующим образом:
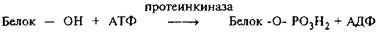
Гидроксильная группа белка, акцептирующая терминальный фосфат АТФ, почти всегда принадлежит серину, реже тАУ треонину и тирозину. Фосфосериновые остатки в белках существенно ионизированы при физиологических значениях рН, поэтому фосфорилирование или дефосфорилирование серина резко меняет заряд белковых молекул. После открытия протеинкиназы А стало ясно, что реакции фосфорилирования тАУ дефосфорилирования опосредуют действие многих гормонов и нейромедиаторов, которые через р-адренергическую рецептию активируют аденилатциклазу и приводят к повышению внутриклеточного уровня цАМФ.
Протеинкиназа А обнаружена во всех нормальных клетках млекопитающих, а также в некоторых типах клеток немлекопитающих. Высоким уровнем протеинкиназы А отличается мозг, где фермент распределен равномерно по всем отделам. Для проявления активности фермента необходимы ионы магния. Так, из легких кролика выделен стимулирующий модулятор протеинкиназы, связывающий магний и названный "магмодулином". Протеинкиназа А тАУ тетрамер, построенный из двух различных субъединиц: регуляторной, связывающей цАМФ, и каталитической, осуществляющей фосфотрансферазную реакцию; будучи соединены вместе, они образуют неактивный комплекс. При активации происходит связывание цАМФ Р-субъединицей холофермента, после чего возможна диссоциация Р от К-субъединицы. Свободная К-рубъединица способна катализировать фосфорилирование различных клеточных белков:
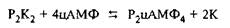
Таким образом, в активном состоянии протеинкиназа А представляет димер Р-субъединиц, связанный с 4 молями цАМФ, и 2 свободные К-субъединицы.
Скорость обратной реакции обусловлена не только спонтанной диссоциацией комплексов Р тАУ цАМФ, но и концентрацией К-субъединиц, так как последние высвобождают связанный цАМФ путем рекомбинации с комплексом Р тАУ цАМФ. Этот своеобразный "ретрокооперативный" эффект К-субъединицы, названный так Свилленсом и Дюмоном, ведет к временному увеличению концентрации цАМФ, требуемой для активации протеинкиназы. В период стимуляции системы уровень К-субъединиц резко падает.
Существование эквивалентных количеств каталитических и регуляторных субъединиц цАМФ-зависимой протеинкиназы во многих тканях привело к представлению о том, что единственная функция регуляторной субъединицы тАУ контроль протеинкиназной активности. Однако регуляторная субъединица может существовать отдельно от каталитической в некоторых типах клеток, например в клетках нейробластомы. Это указывает на возможность проявления биологического действия регуляторной субъединицы независимо от каталитической. Далее будет рассмотрена такая возможность на примере изменения проницаемости мембран нейронов для Na+ и К в присутствии Р-субъединицы. Надо также иметь в виду, что цАМФ-связывающие белки, не ассоциированные с протеинкиназой, могут регулировать активность последней путем модулирования уровня свободного циклического нуклеотида, способного присоединяться к протеинкиназе.
Отметим далее, что свободные Р-субъединицы ингибируют фосфодиэстеразу цАМФ, что может приводить к увеличению уровня этого нуклеотида в клетке. Установлено также, что свободные Р-субъединицы протеинкиназы типа А в отличие от холофермента киназы и ее К-субъединицы ингибируют фосфопротеинфосфатазы нескольких типов. Ингибирование обусловлено снижением скорости катализа без изменения сродства к энзиму. Очевидна физиологическая значимость такого ингибирования, так как в этом случае диссоциация А-киназы может способствовать не только стимулированию фосфорилирующей активности, но и ингибированию дефосфорилирования субстратов.
Протеинкиназа А существует в форме двух изоферментов, относительное количество которых варьирует в разных тканях.
Изоферменты были названы киназами I и II. Они имеют различные скорости диссоциации в присутствии гистона и растворов NaCl и реассоциации после удаления цАМФ. Так, киназа I типа быстро диссоциирует в присутствии гистона или 0,5 М NaCl и медленно реассоциирует после удаления цАМФ; напротив, киназа II типа медленно диссоциирует в присутствии указанных агентов и быстро реассоциирует после удаления цАМФ.
Каталитические субъединицы киназ I и II типа имеют молекулярную массу 40 кД и минимальные различия в аминокислотном составе. Напротив, регуляторные субъединицы киназ I и II типа значительно отличаются по первичной структуре. Вероятно, субъединица Р I типа тАУ 49 кД тАУ является протеолитическим фрагментом Р II типа с молекулярной массой 55 кД. Один из сериновых остатков Р-субъединицы II типа фосфорилируется каталитической субъединицей цАМФ-зависимой протеинкиназы. Очищенная же Р-субъединица I типа не фосфорилируется каталитической субъединицей цАМФ-зависимой протеинкиназы. Другим отличием киназы I типа от II типа является то, что только первый фермент связывает Mg-АТФ с высоким сродством.
цАМФ-зависимые протеинкиназы локализованы в основном в цитозольной фракции клеток. Однако в мозге, например, значительная часть киназы II типа является мембранно-связанной. Очевидно, субклеточная локализация и соотношение киназ I и II типа могут обусловливать специфику действия цАМФ в клетке. Отношение киназы I типа к киназе II типа варьирует в различных органах и в различных фазах клеточного цикла.
При исследовании локализации фосфорилирующих систем в ЦНС установлено, что цАМФ-зависимая система избирательно сконцентрирована в нейронах, особенно в дендритах, а не в глии. Мозг крысы содержит как I, так и и II форму протеинкиназы А при соотношении этих форм 1:4 соответственно. Высокое содержание А-киназы II типа по сравнению с ферментом I типа вообще характерно для нервной ткани. Недавно установлена гетерогенность Р-субъединиц А-киназы II типа. При этом в мозге выявлена собственная, специфичная Р Н-субъединица, отличная по иммунохимическим свойствам от Р-субъединиц II типа в других тканях. Р II мозга отличается от Р II тАУ мышц по характеру взаимодействия с К-субъединицей, а также по электрофоретической подвижности аутофосфорилированных форм. Показано, что фракция Р-субъединиц II типа мозга взаимодействует с Са+ и кальмодулином. Необычные свойства Р Н-субъединицы мозга могут являться следствием адаптации нервных клеток к специфической функции передачи и хранения информации.
Ассоциации А-киназы II типа мозга с мембранной фракцией клеток обусловлены только Р-субъединицей. цАМФ при диссоциации холофермента высвобождает из мембранно-связанного состояния только К-субъединицу; таким образом, компартментализация К-субъединицы изменяется при активации А-киназы. Молекула этой субъединицы гидрофильна, поэтому диссоциация мембранно-связанной А-киназы II типа приводит к транслокации каталитической субъединицы в цитоплазму и далее в ядро. Предполагается, что такая транслокация может обусловливать цАМФ-зависимое изменение экспрессии генов в нейронах, наибольшая концентрация Р-субъединиц А-киназы II типа обнаружена в пре- и постсинаптической мембранах, что свидетельствует о важной роли этой киназы в синаптической передаче.
Различия между регуляторными субъединицами А-киназы II типа нервной и других тканей проявляются также и во взаимодействии этих субъединиц с субстратами киназы. Так, для Р-субъединицы II типа из мозга характерно тесное взаимодействие с МАР-2 тАУ нейроспецифическим белком, локализованным в отростках нейронов, кальцинейрином и другими белками. Такое взаимодействие Р II с субстратами А-киназы может приводить, с одной стороны, к локализации А-киназы II типа около специфических субстратов и, соответственно, в определенных внутриклеточных компартментах, что, очевидно, имеет важное физиологическое значение. С другой стороны, кроме связывания К-субъединицы Р-субъединица II типа нервной ткани может участвовать в регуляции функционирования других белков.
Регуляция активности протеинкиназы А осуществляется несколькими путями. Так, во многих тканях, в том числе и нервной, обнаружен низкомолекулярный термостабильный ингибитор киназы. Ингибитор связывается со свободной К-субъединицей и угнетает ее ферментативную активность. Вероятно, физиологическая роль ингибитора состоит в блокаде фосфорилирующей активности при "базаяном", т.е. нестимулируемом уровне цАМФ. В мозге наблюдается изменение концентрации ингибитора в ответ на некоторые гормональные сигналы, что может иметь определенное значение в долговременной регуляции активности А-киназы. Регуляция активности киназы II типа осуществляется также аутофосфорилированием ее Р-субъединицы; это ведет к повышению активности фермента, что обусловлено уменьшенной скоростью реассоциации фосфорилированной Р со свободной К-субъединицей. Киназа I типа также может быть фосфорилирована, но фосфорилирование ее Р-субъединицы осуществляется цГМФ-зависимой протеинкиназой. Установлено ау-тофосфорилирование К-субъединицы, однако функциональное значение этого процесса неизвестно.
Возможно, что фосфорилирование с помощью свободной К-субъединицы А-киназы и соответствующая активация фосфопротеинфосфатазного ингибитора I, приводящая к снижению активности фосфопротеинфосфатазы I, уменьшает дефосфорилирование комплекса Р-цАЛ1Ф и, следовательно, способствует диссоциации киназы II типа по принципу положительной обратной связи. Такое событие наряду с компартментализацией цАМФ в клетке может способствовать эффективной активации цАМФ-зависимой киназы при незначительном увеличении внутриклеточного уровня цАМФ.
цАМФ-зависимое фосфорилирование белков нервных окончаний оказывает существенное влияние на синаптическую передачу. Так, установлено, что мутации у дрозофил, изменяющие метаболизм цАМФ, приводят к плейотропным нарушениям механизма обучения. У моллюска аплизии серотонинергическая синаптическая передача усиливается инъекцией цАМФ в нейрон или инкубацией ганглия с блокаторами фосфодиэстеразы. Так как известно, что входящий ток кальция является непременным посредником в осуществлении синаптического действия, полагают, что серотонин усиливает такой ток посредством повышения внутриклеточного уровня цАМФ и активности А-киназы. Прямые доказательства осуществления этого механизма получены в экспериментах, где отмечено увеличение Са-проводимости нейронов улитки при инъекции в них К-субъединицы А-киназы. Блокаторы протеинкиназы А оказывают противоположный эффект тАУ быстрое снижение Са-проводимости.
Повышение уровня ионизированного Са+ внутри клетки при открывании Са-каналов может способствовать снижению в ней содержания цАМФ за счет активации Са тАУ КМ-зависимой фосфодиэстеразы и по принципу отрицательной обратной связи приводить к переходу Са-каналов в неактивное состояние. Введение в нейрон теофиллина замедляет процесс ослабления Са-проводимости. Таким же влиянием на кальциевую проводимость обладают ингибиторы кальмодулина, например трифтазин.
Таким образом, функционирование Са-каналов прямо регулируется протеинкиназой А, осуществляющей фосфорилирование их белковых компонентов, а также опосредованно контролируется гормонами, медиаторами и другими факторами, вызывающими или активацию аденилатциклазы, или ее ингибирование. При нарушении фосфорилирования каналы быстро теряют способность активироваться под действием изменения трансмембранного электрического поля. В течение некоторого времени это состояние является обратимым и фосфорилирование снова переводит каналы в рабочее состояние либо изменяя кинетику их активации, либо включая ранее неактивные каналы. При длительном нарушении цАМФ-зависимого фосфорилирования каналы подвергаются необратимым изменениям.
Дополнительный путь для токов Са+ внутрь клетки могут образовывать хемоуправляемые каналы, непосредственно активируемые медиаторными веществами в ответ на их взаимодействие с рецепторами. В отличие от электроуправляемых Са-каналов время жизни этих каналов существенно больше, однако относительное количество ионов, переносимых через хемоуправляемые каналы, вероятно, невелико по сравнению с электроуправляемым путем. По ряду данных хемоуправляемые каналы полностью инактивируются при увеличении внутриклеточного содержания цАМФ.
Заслуживает также рассмотрения участие цАМФ в К- и Na-проводимости мембраны.
Известно, что болевое раздражение усиливает рефлекс втягивания жабры у моллюска эплизии за счет модуляции секреции передатчиков сенсорными нейронами. Повышение чувствительности происходит за счет снижения К-проводимости и соответствующего увеличения продолжительности потенциала действия, в результате чего усиливаются Са-рефлекс и экзоцитоз. Болевое раздражение вызывает выделение серотонина "облегчающим" нейроном в окончаниях сенсорных нейронов. Серотонин, в свою очередь, увеличивает синтез цАМФ, активность протеинкиназы А и степень фосфорилирования белка, тесно связанного с К-каналом. В результате происходит закрытие канала. Блокада К-каналов приводит к тому, что приходящий в нервное окончание потенциал действия спадает медленно: "продленные" потенциалы действия удерживают потенциал-зависимые Са-каналы в открытом состоянии, вследствие чего приток ионов Са возрастает. Это, в свою очередь, ведет к опорожнению большего числа синаптических пузырьков. К блокаде К-каналов приводит также добавление АТФ и К-субъединицы протеинкиназы А в мембранные препараты нейронов; добавление протеинфосфатазы обусловливает открывание К-каналов.
Электрозависимые Na-каналы ответственны за формирование потенциала действия в возбудимых мембранах. Установлено, что в синаптосомах мозга а-субъединица №^каналов, играюшая центральную роль в функционировании этих каналов, фосфорилируется как эндогенной, так и экзогенной протеинкиназой А. Фосфорилирование приводит к ингибированию активированного нейротоксинами входящего Na-тока. Таким образом, активность Са-каналов, К-каналов и Na-каналов нейронов модулируется под действием протеинкиназы А и / или через изменение скорости их дефосфорилирования протеинфосфатазами.
Отметим, что протеинкиназа А в нервной ткани регулирует также чувствительность р-адренорецепторов к агонистам. Так, десенситизация этих рецепторов коррелирует с их цАМФ-зависимым фосфорилированием. Установлена также регуляция А-киназой биосинтеза самих р-агонистов. Эта регуляция осуществляется с помощью цАМФ-зависимого фосфорилирования тирозингидроксилазы тАУ узлового фермента биосинтеза катехоламинов. Такое фосфорилирование может быть составной частью механизма ускорения биосинтеза катехоламинов в ответ на нервный импульс или секрецию нейромедиаторов в нервной ткани в условиях in vivo,
Остановимся на роли цАМФ и еще одного вторичного посредника тАУ 2', 5'-олигоаденилата в регуляции пролиферации и дифференцировки нервных клеток. 2тАУ5А тАУ это олигонуклеотид, состоящий из нескольких остатков АМФ, связанных 5'-фосфодиэфирной связью. В клетке существуют 2 фермента, обеспечивающие определенный уровень 2тАУ5А. Олигосинтетаза тАУ фермент биосинтеза 2тАУ5А, активен лишь в присутствии двуспиральной РНК; молекулярная масса тАУ 100тАУ105 кД. 2'-Фосфодиэстераза гкдролизует 2тАУ5А до АМФ и АТФ, молекулярная масса этого фермента около 40 кД.
Чтобы оценить биохимическое и физиологическое значение процессов образования 2тАУ5А, необходимо остановиться на механизмах подавляющего влияния цАМФ на пролиферативный статус нервных клеток. Так, действие ряда факторов, в частности фактора роста нервов, а также повышение уровня цАМФ, вызванное теофиллином или дибутирил-цАМФ, приводит к остановке деления и дифференцировке этих клеток. Действие теофиллина в отличие от фактора роста нервов или дибутирильного аналога цАМФ носит кратковременный характер. Повышение концентрации цАМФ в результате ингибирования фосфодиэстеразы под действием теофиллина, как и в случае фактора роста нервов и дибутирил-цАМФ, приводит к блокаде активного деления и дифференцировке нервных клеток. Однако в дальнейшем падение активности фосфодиэстеразы, обусловленное теофиллином, и соответствующее увеличение уровня цАМФ компенсируется индукцией биосинтеза этого фермента. Именно этим и объясняется кратковременное действие теофиллина.
Повышение уровня цАМФ в нервных клетках, вызванное действием факторов, непосредственно влияющих на аденилатциклазную систему, сопровождается более чем 10-кратным увеличением концентрации 2тАУ5А. Установлено, что рост уровня олиго синтетазы и концентрации 2тАУ5А наблюдается при дифференцировке клеток и замедлении клеточного деления.
Исследование биологических свойств 2тАУ5А показало, что этот олигоиуклеид способен обратимо активировать специфическую латентную нуклеазу, ускорять гидролиз РНК и, таким образом, ингибировать синтез белка in vivo и тормозить процесс размножения клеток. Следовательно, повышение уровня 2тАУ5А и изменение концентрации цАМФ является частью универсального механизма регуляции клеточного деления. К настоящему времени установлены два взаимодополняющих процесса, с помощью которых осуществляются эти регуляторные реакции. С одной стороны, стимуляция А-киназы при повышении уровня цАМФ приводит к фосфорилированию белка сМг тАУ 18 кД. Этот белок в виде фосфоформы является ингибитором активности 2' тАУ фосфодиэстеразы тАУ фермента гидролиза 2тАУ5А. С другой стороны, повышение концентрации цАМФ внутри клетки вызывает индукцию олиго синтетазы. осуществляющей синтез 2тАУ5А.
Совокупность этих событий приводит к устойчивому повышению уровня 2тАУ5А и связанному с этим переходу нервных клеток в состояние покоя. По-видимому, именно таким образом осуществляется ярко выраженный антинролиферативный эффект цАМФ, вызванный действием этого вторичного посредника на систему метаболизма 2тАУ5А. При этом взаимное влияние уровней цАМФ и 2тАУ5А основано на механизме отрицательной обратной связи. Так, установлено, что подъем уровня 2тАУ5А, обусловленный увеличением внутриклеточной концентрации цАМФ по указанным механизмам, в свою очередь, способствует активации фосфодиэстеразы цАМФ. В результате происходит снижение уровня цАМФ и прекращение роста концентрации 2тАУ5А, который является в этом случае вторичным посредником в проявлении антипролифера-тивного действия цАМФ.
1.3 Фосфодиэстераза
В прекращении сигнала цАМФ участвуют фосфодиэстераза, гидролизующая этот нуклеотид до АМФ. В противоположность аденилатциклазе фосфодиэстераза тАУ преимущественно растворимый фермент. В то же время активность фермента обнаружена во фракциях саркоплазматического ретикулума, митохондрий и ядер.
Циклонуклеотидактивируемая фосфодиэстераза гидролизует как цАМФ, так и цГМФ, при этом под действием цГМФ ускоряется гидролиз цАМФ и наоборот, что свидетельствует о положительной кооперативности двух активных центров. Несомненна роль фермента в проведении нервного импульса: так, в постсинаптической мембране нервной ткани обнаружен необычайно высокий уровень фосфодиэстеразы цГМФ.
Для фосфодиэстеразы характерно наличие множественных форм. Эти формы различаются как по молекулярной массе, так и по сродству к одному и тому же и к различным циклическим нуклеотидам. В целом, сродство к циклическим нуклеотидам у фосфодиэстераз в 100тАУ1000 раз ниже, чем у протеинкиназ А и G, поэтому при ускорении синтеза нуклеотидов сначала происходит насыщение циклическими нуклеотидами регуляторных центров киназ и лишь затем тАУ гидролиз цАМФ и цГМФ фосфодиэстеразами.
В мозге содержатся Са-кальмодулинрегулируемые формы как цАМФ-синтезирующего, так и цАМФ-гидролизующего ферментов. Са-кальмодулинзависимая фосфодиэстераза представляет собой гомодимер, состоящий из субъединиц с различной молекулярной массой у разных изоформ фермента. Ограниченный протеолиз субъединицы 60 кД приводит к появлению фрагмента с Мг = 36 кД и необратимой активации фосфодиэстеразы. Фрагмент 36 кД более не активируется кальмодулином. Следовательно, субъединицы фосфодиэстеразы включают 2 фрагмента: каталитический и регуляторный.
Анализ регуляторных свойств фосфодиэстеразы в нервной ткани свидетельствует о тесном сопряжении между цАМФ- и Са-зависимыми системами внутриклеточной сигнализации; это сопряжение может модулироваться с помощью изоферментов фосфодиэстеразы. Так, в мозге быка найдены 2 изоформы Са-КМ-зависимой фосфодиэстеразы, состоящей из субъединиц с Мг = 60 и 63 кД. Изофермент с субъединицами 60 кД может быть фосфорилирован цАМФ-зависимой протеинкиназой, что приводит к уменьшению сродства фосфодиэстеразы к кальмодулину. Дефосфорилирование этого изофермента осуществляет Са-КМ-стимулируемая протеинфосфатаза; при этом восстанавливается чувствительность фосфодиэстеразы к кальмодулину.
В отличие от изофермента с субъединицами 60 кД фосфорилирование изоформы фосфодиэстеразы с субъединицами 63 кД осуществляется Са-КМ-зависимыми протеинкиназами. Это фосфорилирование также приводит к потере чувствительности фосфодиэстеразы к кальмодулину и "обращается" Са-КМ-стимулируемой протеинфосфатазой с восстановлением чувствительности фосфодиэстеразы к КМ. Очевидно, такой механизм регуляции фосфодиэстеразы реализуется в мозге in vivo, несмотря на очень высокую, "насыщающую" концентрацию в нем КМ. Фосфорирование снижает сродство фермента к КМ и обусловливает зависимость его активности от физиологических концентраций Са+.
В нервной ткани, таким образом, существует тесная взаимосвязь между двумя системами вторичных посредников, Са+ и цАМФ, осуществляемая посредством цАМФ- и Са-зависимого фосфорилирования-дефосфорилирования различных изоферментов фосфодиэстеразы цАМФ. Эта взаимосвязь может модулироваться также различным сродством к Са+ аденилатциклазы, фосфодиэстеразы, протеинкиназы и фосфатазы. Так, аденилатциклаза мозга активируется гораздо более низкими концентрациями Са чем фосфодиэстераза.
Кроме циклических нуклеотидов, Са+ и ограниченного протеолиза фосфодиэстеразу активируют также полианионы, фосфолипиды, жирные кислоты. Ингибиторами или активаторами фермента являются многочисленные фармакологические вещества. Фосфодиэстераза является более "удобной" мишенью для действия лекарственных препаратов, чем аденилатциклаза, так как менее специфична в отношении эффекторных влияний. Мощными ингибиторами фосфодиэстеразы являются производные ксантинов тАУ ингибирование осуществляется блокадой аллостерического центра связывание нуклеотидов.
2. цГМФ-зависимое протеинфосфорилирование
Вскоре после открытия цАМФ-зависимых протенкиназ был обнаружен еще один класс циклонуклеотидзависимых фосфорилирующих ферментов, стимулируемых с помощью цГМФ, тАУ протеинкиназы G. В тканях млекопитающих содержание протеинкиназ G весьма невелико. Наиболее высок уровень активности и содержания протеинкиназ G в мозжечке, сердечной мышце и легких, эти же ткани содержат и наибольшее количество цГМФ, не превышающее, впрочем, 10% от содержания в них цАМФ. Основное количество протеинкиназы G обнаружено в цитозоле, некоторая часть фермента связана с цитоплазматическими мембранами и ядерной фракцией.
Фермент состоит из двух примерно идентичных субъединиц, каждая из которых имеет каталитическую активность и способна связывать циклический нуклеотид. Ограниченным протеолизом можно перевести димер в мономеры и затем разделить каждую субъединицу на цГМФ-связывающий и каталитический фрагменты. Эти исследования наряду с выявленной гомологией между протеинкиназой G и протеинкиназой А II типа по субстратной специфичности, аминокислотной последовательности, способности к аутофосфорилированию, иммунологическим свойствам привели к представлению о том, что цАМФ и цГМФ-зависимые протеинкиназы эволюционировали от общего гена, кодирующего одну полипептидную цепь. В процессе развития цАМФ-зависимый фермент стал синтезироваться в виде разобщенных компонентов, тогда как цГМФ-зависимая протеинкиназа синтезируется как одна цепь.
В отличие от каталитической субъединицы протеинкиназы А, способной высвобождаться во время активации фермента, внутриклеточное перемещение "недиссоциированной" протеинкиназы G может быть затруднен. На основании аминокислотного анализа протеинкиназы G было установлено, что одна субъединица фермента содержит два центра связывания нуклеотида.
Регуляторная N-концевая половина молекулы протеинкиназы G сходна с семейством цАМФ-связывающих белков, в то время как каталитическая Сконцевая половина родственна группе киназ с различной специфичностью. Протеинкиназа G способна к аутофосфорилированию из расчета 2 моля фосфата на 1 моль холофермента. Аутофосфорилирование не изменяет сродства к цГМФ и незначительно повышает Vmakc фосфотрансферазной реакции, но заметно увеличивает сродство протеинкиназы G к цАМФ. Таким образом, аутофосфорилирование может обусловливать регуляцию протеинкиназы G не только с помощью цГМФ, но и цАМФ.
Протеинкиназа G фосфорилирует, вероятно, те же аминокислотные остатки в молекуле субстрата, что и протеинкиназа А, но с гораздо меньшей скоростью. Различная скорость фосфорилирования протеинкиназ А и G может являться основой их субстратной специфичности.
Как упоминалось, наибольшее содержание протеинкиназы G в нервной ткани отмечено в мозжечке. В свою очередь, в этом отделе мозга фермент локализован в клетках Пуркинье. Найдена корреляция между увеличением содержания G-киназ в цитоплазме клеток Пуркинье, началом роста дендритов и установлением синаптических контактов в клетках Пуркинье и подобных им субкортикальных клетках, что может свидетельствовать в пользу безусловной значимости цГМФ-зависимого фосфорилирования для нейрональной дифференцировки этих клеток. В клетках Пуркинье находится и единственный в мозге млекопитающих специфичный субстрат для протеинкиназы G, названный G-субстратом; G-субстрат тАУ кислоторастворимый и термостабильный белок с Мг = 23 кД. Обнаружено, что фосфорилированный с помощью цГМФ-зависимой протеинкиназы G-субстрат ингибирует протеинфосфатазу, выделенную из мозжечка, являясь специфичным для клеток Пуркинье протеинфосфатазным ингибитором. При этом ингибируемая фосфатаза из этих клеток, вероятно, катализирует дефосфорилирование белков, не являющихся субстратами протеинкиназы А.
Увеличение уровня цГМФ обусловлено взаимодействием с плазмалеммой различных гормонов и нейромедиаторов. Так активируется связанная с мускариновыми рецепторами гуанилатциклаза Активация гуанилатциклазы обычно обусловлена мобилизацией Са+ из эндоплазматического ретикулума.
В клетках мозга обнаружен термостабильный белок, стимулирующий активность протеинкиназы G, но не изменяющий активность протеинкиназы А. Обратный процесс тАУ инактивация протеинкиназы G состоит в гидролизе цГМФ фосфодиэстеразой, специфичной для этого нуклеотида. Кроме того, в тканях млекопитающих, в том числе и нервной, найден ингибитор G-киназы с Мг = 15 кД.
Отметим, что системы циклических нуклеотидов осуществляют внутриклеточную регуляцию в тесном взаимодействии друг с другом. Так, например, если концентрация цАМФ в клетке длительное время повышена, может происходить фосфорилирование белков, встроенных в каналы плазматической мембраны, что приводит к повышению в цитоплазме концентрации Са+. В результате этого активируются фосфодиэстераза и гуанилатциклаза; соответственно ускоряется гидролиз цАМФ и образуется цГМФ.
Итак, цГМФ играет важную роль в нервной системе, особенно в клетках Пуркинье мозжечка. Об этом свидетельствует избирательная локализация в этих клетках всех компонентов системы цГМФ, включая гуанилатциклазу, протеинкиназу G и G-субстрат. Различные нейромедиаторы, в том числе ацетилхолин, вызывают увеличение уровня цГМФ в клетках Пуркинье и активацию G-киназы, что, по всей видимости, модулирует такие свойства этих клеток, как скорость проведения возбуждения и способность к стимуляции.
Установлено также, что цГМФ модулирует активность ионных каналов мембран нервных клеток. Так, введение в нейроны улитки цГМФ или G-киназы увеличивает проводимость Са-каналов.
3. Са-кальмодулинзависимое протеинфосфорилирование
Как упоминалось, в процессе эволюции выработались эффективные механизмы удаления Са+ во внеклеточное пространство или специализированные внутриклеточные структуры. Низкая внутриклеточная концентрация Са+ позволила клеткам использовать "впрыскивание" Са+ в цитоплазму как сигнал на внешние воздействия. При этом кальций может быть даже более универсальным внутриклеточным регулятором, чем цЛМФ. Так, период действия Са+ очень короток и измеряется миллисекундами, а не секундами, как в случае цАМФ. Как стало понятно в последнее время, обе системы вторичных посредников функционируют в тесном взаимодействии.
Для внутриклеточной "реализации" кальциевого сигнала в процессе эволюции созданы специальные внутриклеточные белковые рецепторы Са+, способные опосредовать действие этого иона на молекулярные мишени. Основным рецептором Са+ во всех клетках является кальмодулин. Кальмодулин тАУ глобулярный белок сМг тАУ 16,5 кД. Структурно он очень консервативен: так, обнаружено только шесть или немногим более аминокислотных замен в кальмодулине, выделенном из живых объектов, эволюционно разделенных миллионами лет. Кальмодулин содержит 4 Са-связывающих участка: в состав каждого из этих участков входят кислые остатки аминокислот, необходимые для связывания Са+
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)