Работа электрических органов рыб
Электрическое оружие и электролокаторы
На электрических рыбах впервые достоверно установили возможность выработки электричества животными, на них Фарадей показал, что электричество животных ничем не отличается от электричества гальванических элементов или электрических генераторов. Как же устроены электрические органы рыб?
Основу этих органов составляют столбики из плоских клеток, лежащих друг на друге, как пары медь тАУ цинк в вольтовом столбе или как стопка монет, К одной поверхности каждой клетки подходит нервное окончание. Когда орган находится в покое, обе стороны каждой клетки имеют одинаковый потенциал и ток через орган не идет, Когда же по всем нервным волокнам одновременно приходят импульсы, постсинаптическая мембрана резко повышает свою проницаемость к ионам и потенциал на ней падает до нуля. Это приводит к возникновению тока, текущего через клетку. Все клетки столбика соединены последовательно и поэтому их потенциалы суммируются, как в последовательно соединенных гальванических элементах. Такое объяснение дал работе электрических органов рыб создатель мембранной теории биопотенциалов
Ю. Бернштейн в своей книге ВлЭлектробиологияВ». Это объяснение в своей основе оказалось верным, а для некоторых электрических рыб и вполне точным. Что касается электрического угря, то у него мембрана той стороны клетки, на которую действует синапс, оказалась электрически возбудимой, так что при приходе нервного импульса она не просто снижает свой потенциал до нуля, а перезаряжается, что обеспечивает более высокую разность потенциалов, генерируемую каждой клеткой. У скатов и звездочетов клетки электрических органов таковы, что ни одна из их мембран не способна генерировать импульсы.
Читатель уже знает, что одиночная клетка обладает ПП порядка 60 мВ и даже при возбуждении возникающий ПД имеет амплитуду всего порядка 120 мВ. Между тем*' электрический угорь умеет создавать напряжение 800 тАУ 900 В, а нильская щука и нильский сом тАУ напряжение 200тАУ350 В. Как говорилось, это обеспечивается последовательным соединением многих клеток. У электрического угря последовательно соединены более 6 000 клеток,
Однако задача электрического органа состоит не в том, чтобы создать возможно большую Э.Д.С. Дело в том, что часть Э.Д.С. всегда ВлтеряетсяВ» на внутреннем сопротивлении источника. А электрический орган должен обеспечивать возможно большую разность потенциалов в наружной среде. Ясно, что чем большую силу тока обеспечит электрический орган, тем больше будет и падение напряжения в наружной среде. Рассмотрим эту проблему более подробно.
Сила тока, создаваемого электрическими органами и текущего через воду, по закону Ома для всей цепи равна: / = E/. Если у нас имеется много гальванических элементов, то их можно соединить либо последовательно, либо параллельно, либо смешанным образом, образуя ВлстолбикиВ» из последовательно соединенных элементов и соединяя эти ВлстолбикиВ» параллельно друг другу. Пусть в нашем распоряжении имеется ограниченное число элементов тАУ п. Пусть сопротивление наружной среды задано и равно R, а внутреннее сопротивление элемента равно г и его э. д. с. равна Е. Как выгоднее соединить элементы, чтобы получить максимальную силу тока в цепи, а значит и максимальное падение напряжения на внешнем сопротивлении? Давайте будем соединять в один столбик х элементов, тогда число столбиков будет равно п/х. Общая э. д. с. такой системы будет равна Ех. Внутреннее сопротивление одного столбика будет равно гх, а полное внутреннее сопротивление всей системы в nix раз меньше, т.е. равно гх2/п. Тогда для силы тока мы получим выражение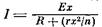 , Это выражение имеет максимум, когда гхг1п = R, т.е. когда внутреннее сопротивление всей системы равно внешнему.
, Это выражение имеет максимум, когда гхг1п = R, т.е. когда внутреннее сопротивление всей системы равно внешнему.
А теперь посмотрим, в какой мере законы физики влияли на ход эволюции электрического органа. Для пресноводных рыб сопротивление окружающей среды весьма велико и для достижения максимального эффекта необходимо увеличивать х, т.е. число последовательно соединенных элементов. Поэтому электрический угорь тАУ обитатель болот Южной Америки тАУ имеет электрический орган, клетки которого в основном соединены последовательно, в то время как столбиков у него мало. Совсем другая ситуация у ската Торпедо, обитателя Средиземного моря, ведь сопротивление соленой морской воды относительно мало, И в электрическом органе ската клетки соединены совсем иначе: последовательно соединены всего 400 клеток, зато число параллельно соединенных столбиков велико. В результате электрический орган ската создает силу тока порядка 50 А, так что падение напряжения даже в соленой воде оказывается достаточно заметным, порядка 50 В, Таким образом, общий ход эволюции электрических органов вполне соответствует ожиданиям физика.
Мы видим, что морским электрическим рыбам важно сделать внутреннее сопротивление своего генератора как можно ниже, для этого они и соединяют так много столбиков параллельно. Но есть и еще один способ снизить это сопротивление тАУ снизить сопротивление мембран каждой клетки. Оказывается, эволюция использует и этот способ. У одной мембраны клеток органа, той, на которой находится синапс, сопротивление при возбуждении становится низким, так как в ней открываются многочисленные ионные каналы, а сопротивление противоположной мембраны тоже относительно невелико за счет ее большой площади: она вся покрыта складками, выростами и т.д.
Законы физики и условия среды обитания определяют пути эволюции электрических органов, и последние развиваются сходным образом, хотя и происходят у разных рыб из разных тканей. Например, у скатов клетки электрических органов возникли из мышечных клеток, потерявших возбудимость, фактически, от этих клеток осталась только синаптическая область; постепенная атрофия остальной части мышечной клетки хорошо прослежена при изучении развития рыбы из икринки. Медиатор, действующий на электрический орган, тАУ тот же ацетилхолин, который возбуждает и мышечные клетки позвоночных, поэтому разряд органа можно вызвать не только раздражением подходящего к нему нерва, но и введением, ацетилхолина в идущую к нему артерию. У электрического угря орган тоже имеет мышечное происхождение, однако у некоторых электрических рыб электрические органы возникли не из мышечных, а из нервных клеток, а у африканского электрического сома тАУ из клеток кожных желез. Это не должно удивлять читателя, который уже знает, что не только нервы и мышцы способны к электрическим реакциям.
Такие рыбы, как электрический скат или электрический угорь, затрачивают при разрядах электрического органа заметную энергию, В связи с этим они используют свои электрические органы сравнительно редко. У электрического угря есть еще дополнительные электрические органы, гораздо меньшей мощности, которые используются для ориентации и обнаружения добычи. У слабоэлектрических рыб, которые используют свои электрические органы только для ориентации, часто наблюдается совсем другой режим работы тАУ непрерывные разряды. Например, у рыбы гимнарха электрические органы постоянно работали с частотой 300 Гц.
Важная проблема, которая стоит перед всеми электрическими рыбами, тАУ проблема синхронизации, т.е. проблема вызова одновременного возбуждения всех клеток, образующих электрический орган. Она решается, прежде всего, с помощью ЭС. Нейроны разных уровней, управляющие электрическим органом, связаны между собой ЭС и поэтому разряжаются практически одновременно. У электрических рыб обнаружены рефлекторные цепи, которые ведут к возбуждению электрического органа; в этих цепях осуществляется последовательная передача сигналов между тремя типами нейронов, причем сигналы передаются только через ЭС. Однако одновременного возбуждения нейронов, действующих на клетки электрического органа, еще не достаточно для одновременного возбуждения самих этих клеток, так как сотни и тысячи клеток ВлстолбиковВ» расположены на разных расстояниях от нейронов. Аксоны этих нейронов имеют разную скорость проведения: к более удаленным клеткам сигнал идет с большей скоростью.
Как поймать рыбу в мутной воде? А также про электроразговоры
Для существа, обладающего чувствительными электрорецепторами, обнаружить рыбу в мутной воде несложно; для этого достаточно зарегистрировать электрические поля, возникающие при работе ее сердца или дыхательных мышц. Так обнаруживают добычу акулы или скаты. Но рыбы, обладающие электрическими органами, оказались способными решить и более хитрую задачу: обнаружить в непрозрачной воде предметы, которые сами по себе не создают электрических полей. Как же электрические рыбы решают эту задачу?
Сначала думали, что эти рыбы обладают электролокаторами, такими же, как радиолокаторы для обнаружения самолетов. Предполагалось, что эти рыбы генерируют своими электрическими органами сигнал и принимают отраженную от посторонних предметов волну с помощью электрорецепторов. Однако простейшая прикидка показывает, что такой принцип работы невозможен для живых организмов. Рыба обнаруживает посторонние предметы на расстоянии в десятки сантиметров, а электромагнитные волны проходят такие расстояния за слишком короткие времена. Миллиардные доли секунды живые существа измерять не могут. Как же рыбы с помощью электрических органов обнаруживают в воде посторонние предметы?
Лиссман изучал эту способность рыб так. У рыбы вырабатывался условный рефлекс. В аквариум, в котором она содержалась, помещали два цилиндрика одинаковых размеров, обладающих таким же удельным сопротивлением, как вода аквариума, и пустых внутри. В цилиндрики помещались вещества с разным удельным сопротивлением. Рыбу обучали выбирать цилиндрик с более низким удельным сопротивлением. При правильном выборе она получала пищу, а при неправильном тАУ удар палочкой. Через некоторое время рыба уверенно выбирала нужный цилиндрик, несмотря на то, что два цилиндрика все время меняли местами случайным образом. Если же в цилиндриках находились вещества, отличающиеся химическим составом, плотностью и др., но с одинаковым удельным сопротивлением, обучение было безуспешным. Итак, рыбы при обнаружении посторонних предметов могут использовать отличие их удельного сопротивления от сопротивления воды.
Мы уже говорили, что гимнарх все время генерирует электрические импульсы, так что вокруг его тела в воде текут токи. Если изобразить плотность тока с помощью густоты линий, то картина токов вокруг рыбы выглядит примерно так, как изображено на рис. 64. Если в воду внесено проводящее тело, то картина меняется, как на рис. 64, справа, если изолятортАУкак на рис. 64, слева. Искажение электрического поля меняет сигналы электрорецепторов рыбы.
Нервная система электрической рыбы содержит сложную систему обнаружения. Ее мозг сравнивает сигналы от многих рецепторов. Это позволяет определить размеры, форму, скорость движения лоцируемого предмета. Можно сказать, что электрические рыбы обладают настоящим Влэлектрическим зрениемВ».
Электрические рыбы используют электрические сигналы и для общения между собой. Например, они оповещают других особей своего вида о том, что данная территория занята, или о том, что ими обнаружена пища; есть электрические сигналы: Влвызываю на бойВ», ВлсдаюсьВ» и др. По-видимому, электрические сигналы облегчают рыбам поиск особей другого пола. Все эти сигналы эффективно принимаются рыбами на расстоянии порядка 10 метров,
Что такое ЭК-Г, ЭМГ, ЭЭГ?
ЭКГ тАУ это электрокардиограмма, запись электрических сигналов сердца. То, что в сердце при возбуждении возникает разность потенциалов, было показано еще в 1856 г., в эпоху Дюбуа-Реймона. Опыт, доказывающий это, был поставлен Келликером и Мюллером точно по рецепту Гальвани: на изолированное сердце клался нерв, идущий к лапке лягушки, и этот Влживой вольтметрВ» отвечал вздрагиванием лапки на каждое сокращение сердца.
С появлением чувствительных электроизмерительных приборов стало возможным улавливать электрические сигналы работающего сердца, прикладывая электроды не прямо к сердечной мышцех а к коже. Сто лет назад, в 1887 r.j впервые удалось зарегистрировать таким способом ЭКГ человека, Это было сделано английским ученым А. Уоллером с помощью капиллярного электрометраВ» Этот прибор был неудобен в использовании и широкое применение электрокардиографии началось позже, после появления в 1903 г. более совершенного прибора тАУ струнного гальванометра Эйнтховена. После появления этого прибора в ряде лабораторий начали детально изучать, чем отличается ЭКГ здорового сердца и сердца при разных заболеваниях. За эти работы В. Эйнтховен получил в 1924 г. Нобелевскую премию, а советский ученый А.Ф. Самойлов, много сделавший для развития электрокардиографии, получил в 1930 г. Ленинскую премию. В результате следующего шага в развитии техники электрокардиографы стали использоваться в каждой крупной больнице.
Какова природа ЭКГ? При возбуждении любого нервного или мышечного волокна ток в одних его участках втекает через мембрану внутрь волокна, а в других тАУ вытекает наружу. При этом ток обязательно течет по наружной среде, окружающей волокно, и создает в этой среде разность потенциалов. Это позволяет регистрировать возбуждение волокна с помощью внеклеточных электродов, не проникая внутрь клетки. Сердце тАУ это достаточно мощная мышца. В ней синхронно возбуждается много волокон, и в среде, окружающей сердце, течет достаточно сильный ток, который даже на поверхности тела создает разности потенциалов порядка 1 мВ. Обычная форма ЭКГ приведена на рис. 65; зубец Р соответствует возбуждению предсердий, а самый большой зубец R тАУ синхронному возбуждению желудочков.
Для того чтобы больше узнать по ЭКГ о состоянии сердца врачи записывают много кривых между разными точками тела, Для понимания этих кривых нужен большой опыт. С появлением вычислительной техники стало возможным в значительной мере автоматизировать процесс ВлчтенияВ» ЭКГ, Вычислительная машина сравнивает ЭКГ данного больного с образцами, хранящимися в ее памяти, и выдает врачу предполагаемый диагноз. Сейчас возникло много и других новых подходов к анализу ЭКГ. Очень интересным представляется такой. По потенцаалам, зарегистрированным с многих точек тела, и их изменению во времени можно рассчитать, как движется волна возбуждения по сердцу и какие участки сердца стали невозбудимы. Расчеты эти весьма трудоемки, но они стали возможны с появлением ЭВМ. Такой подход к анализу ЭКГ был развит сотрудником Института проблем передачи информации АН СССР Л.И. Титомиром, Вместо многих кривых, в которых трудно разобраться, вычислительная машина рисует на экране сердце и распространение возбуждения по его отделам. Можно прямо видеть, в какой области сердца возбуждение идет медленнее, какие участки сердца вообще не возбуждаются и т.д.
Потенциалы сердца были использованы в медицине не только для диагностики, но и для управления медицинской аппаратурой. Представьте себе, что врачу необходимо сделать рентгеновские снимки сердца в разные фазы его цикла, т.е. в момент максимального сокращения, максимального расслабления и т.д.; это бывает необходимо при некоторых заболеваниях. Но как поймать момент наибольшего сокращения? Приходится делать много снимков в надежде, что один из них попадет в нужную фазу. И вот советские ученые В.С. Гурфинкель, В.Б. Малкин и М.Л. Цетлин решили включать рентгеновскую аппаратуру от зубца ЭКГ, Для этого потребовалось не очень сложное электронное устройство, которое включало съемку с заданной задержкой относительно зубца ЭКГ, Остроумное само по себе решение задачи особенно интересно тем, что это было одно из первых устройств, в которых естественные потенциалы организма управляют теми или иными искусственными устройствами; эта область техники получила название биоуправления.
Скелетные мышцы тела тоже генерируют потенциалы, которые можно регистрировать с поверхности кожи. Однако для этого требуется более совершенная аппаратура, чем для регистрации ЭКГ. Отдельные мышечные волокна обычно работают асинхронно, их сигналы, накладываясь друг на друга, частично компенсируются, и в результате получаются меньшие потенциалы, чем в случае ЭКГ. Электрическая активность скелетной мышцы называется электромиограммой тАУ ЭМГ. Впервые ПД мышечных волокон человека обнаружил, прослушивая их с помощью телефонного аппарата, русский ученый Н.Е. Введенский еще в 1882 г. В 1907 г. немецкий ученый Г. Пипер использовал для их объективной регистрации струнный гальванометр. Однако это был сложный и трудоемкий метод. Только после того как в 1923 г. появился катодный осциллограф и электронная техника, электромиография стала усиленно развиваться. Сейчас ее широко применяют в науке, в медицине, в спорте, а также для биоуправления. Одно из первых замечательных применений биоуправления с помощью ЭМГ тАУ создание протезов для людей, потерявших руку. Такие протезы впервые были созданы в нашей стране.
А что такое ЭЭГ? Это электроэнцефалограмма, т.е. электрическая активность мозга, колебания потенциала, создаваемые работой нейронов мозга и регистрируемые прямо с поверхности головы. Нервные клетки, как и мышечные волокна, работают неодновременно: когда одни из них создают на поверхности кожи положительный потенциал, другие создают отрицательный. Взаимная компенсация потенциалов тут еще сильнее, чем в случае ЭМГ. В результате амплитуда ЭЭГ примерно в сто раз меньше, чем ЭКГ, поэтому их регистрация требует более чувствительной аппаратуры. Впервые ЭЭГ была зарегистрирована русским ученым В, В. Правдич-Немским на собаках с помощью струнного гальванометра; он вводил собакам кураре, чтобы более сильные мышечные токи не мешали регистрации токов мозга. В 1924 г. немецкий психиатр Г. Бергер начал в Йенском университете изучение ЭЭГ человека. Он описал периодические колебания потенциалов мозга, имеющие частоту около 10 Гц, которые называют альфа-ритмом, Он же впервые зарегистрировал ЭЭГ человека при припадке эпилепсии и пришел к выводу, что Гальвани был прав, предполагая, что при эпилепсии в нервной системе возникает участок, где токи особенно сильны. Поскольку речь шла об очень слабых потенциалах, зарегистрированных малоизвестным врачом, результаты Бергера долго не привлекали внимания; он сам опубликовал их только через 5 лет после сделанного открытия. II только после того как в 1930 г. их подтвердили знаменитые английские ученые Эдриан и Мэтьюс, на них была ВлтАжпоставлена печать академического одобренияВ», по выражению Г. Уолтера, английского ученого, который занимался клиническими аспектами ЭЭГ в лаборатории Голла. В этой лаборатории были разработаны методы, которые позволяли по ЭЭГ определять местоположение опухоли или кровоизлияния в мозгу, подобно тому, как ранее научились по ЭКГ определять место инфаркта в сердце. В дальнейшем кроме альфа-ритма были открыты и другие ритмы мозга, в частности ритмы, связанные с разными типами сна. Существует масса проектов биоуправления с помощью ЭЭГ. Например, если у водителя все время регистрировать ЭЭГ, то можно с помощью ЭВМ определить момент, кода он начинает дремать, и будить его. К сожалению, все такие проекты пока трудно реализовать, так как амплитуда ЭЭГ очень мала.
Кроме ЭЭГ тАУ колебаний потенциала мозга в отсутствие специальных воздействий, существует еще и другая форма потенциалов мозга тАУ вызванные потенциалы. ВП тАУ это электрические реакции, возникающие в ответ на вспышку света, звук и т.д. Так как на яркую вспышку света отвечают почти одновременно сразу много нейронов мозга, то ВП обычно имеют гораздо большую величину, чем ЭЭГ. Не случайно они были обнаружены гораздо раньше, чем ЭЭГ. С помощью ВП можно решать интересные научные задачи. Например, после вспышки света ответ раньше всего возникает в затылочной области мозга. Отсюда можно сделать вывод, что именно в эту область поступают сигналы о свете. При электрическом раздражении кожи ВП возникают в темной области мозга. При раздражении кожи руки они возникают в одном месте, кожи ноги тАУ в другом. Можно составить карту таких ответов и эта карта показывает, что поверхность кожи дает проекцию на теменную область коры мозга человека. Интересно, что при этом проектировании нарушаются некоторые пропорции, например проекция кисти руки оказывается непропорционально большой. Да это и естественно: о руке мозгу нужна гораздо более подробная информация, чем, например, о спине,
Электрическое хозяйство инфузории
В этой книге мы рассказывали вам об электрических явлениях в клетках самых разных животных тАУ лягушки и человека, усоногого рака и кальмара. Но все это были многоклеточные животные. А сейчас обратимся к простейшим. Простейшие замечательны тем, что это одноклеточные животные, т.е., с одной стороны, это всего одна клетка, а с другой тАУ организм с достаточно сложным поведением. Этим обусловлены замечательные особенности простейших. У многоклеточных существует разделение труда между разными типами клеток, у простейших, напротив, все функции совмещены в пределах одной клетки, могут выполняться только ею и должны быть согласованы между собой.
Посмотрим же, как живет известная всем со школьных времен тАУ с 6-го класса тАУ пресноводная инфузория туфелька и какую роль в ее жизни играют электрические процессы.
Значительная часть тела туфельки покрыта прочным панцирем из правильно расположенных шестиугольников, в котором имеются ряд просветов для рта, места выброса остатков пищи и др. Кроме того, на поверхности тела туфельки находятся около 15 тысяч ресничек тАУ ее органов движения. Наконец, на поверхности находятся органы защиты тАУ трихоцисты, устройства, которые при раздражении выбрасывают наружу длинную нить, выделяющую ядовитое вещество так же как стрекательные клетки крапивы.
Некоторые реснички, расположенные около рта инфузории, служат не для движения, а для того чтобы загонять пищу в рот. У туфельки рот все время открыт, и она непрерывно питается бактериями. У некоторых других хищных инфузорий рот открывается в момент захвата пищи. Пища, попавшая в рот, через глотку проходит в мембранный пузырек, который отрывается от глотки и совершает сложное путешествие внутри тела инфузории. Этот пузырек называют пищеварительной вакуолью. Таких вакуолей в один и тот же момент может быть много: одни только оторвались от глотки, другие прошли часть пути, третьи уже подходят к специальному участку поверхности, где выбрасываются наружу непереваренные остатки пищи. С нашей, человеческой точки зрения, это довольно необычная пищеварительная система: вместо того чтобы пища двигалась по кишечнику, как у всех людей, у инфузории нет никакого аналога кишечника, а сам ВлжелудокВ» с пищей отправляется в путешествие по телу. Примерно каждые две минуты у туфельки образуется новая пищеварительная вакуоль.
У инфузории есть специальный орган для регуляции содержания воды в теле тАУ выделительная вакуоль. Рассказывая про ионные насосы, мы уже говорили, что у всех пресноводных животных возникает одна и та же проблема: вода за счет осмоса должна поступать внутрь клеток таких организмов и ее излишки необходимо все время удалять. Ту же проблему приходится решать и инфузории. Хотя часть ее тела и защищена плотным покровом, видимо, непроницаемым для воды, но через постоянно открытый рот вода вместе с добычей поступает в организм туфельки. Для выведения лишней воды и служит выделительная вакуоль; такие вакуоли имеются у большинства пресноводных простейших. У туфельки вакуоль состоит из центрального резервуара, подходящих к нему изнутри тела пяти-семи мембранных трубочек и вынодящего канала, который ведет в наружную среду. Удаляемая жидкость собирается в приводящие каналы; в некоторый момент все эти каналы разом сокращаются, растягивая выделительную вакуоль. После этого вакуоль сокращается и выбрасывает жидкость наружу. Вакуоль сокращается примерно один раз в 20 секунд. За 45 минут вакуолью выделяется объем воды, равный всему объему туфельки!
Таким образом, сократительная вакуоль совершает очень большую работу; значительную часть своей энергии туфелька тратит на борьбу с осмосом.
А теперь рассмотрим некоторые электрические явления у инфузории и их связь с ее жизнедеятельностью и поведением. У туфельки существует ГШ; показано, что, как и у других клеток, этот ПП обеспечивается ионами калия. Однако при регистрации этого потенциала оказалось, что он очень неустойчив, У инфузории МП раз в 20 секунд делает очень большой выброс, да и в промежутках сильно колеблется. Оказалось, что эти колебания связаны с разнообразными проявлениями жизнедеятельности туфельки. Когда сократительная вакуоль выбрасывает году и раздувает выводящий канал, она сильно сообщается с наружной средой. Сопротивление мембраны вакуоли ш же, чем других участков тела инфузории, и потенциал на ней тоже ниже, поэтому в момент выброса воды эта вакуоль ВлзакорачиваетВ» остальную мембрану и ПП заметно снижается. То же самое происходит при разрыве пищеварительной вакуоли, когда она сливается с наружной мембраной, а у многих инфузорий тАУ и при открывании рта. Кроме того, у инфузорий имеются разнообразные Влорганы чувствВ», которые тоже все время меняют их МП.
В протоплазме инфузорий гораздо больше калия и гораздо меньше натрия, чем в окружающей воде. Это значит, что туфелька имеет где-то в своем хозяйстве Ка тАУ К-насос. Оказалось что концентрация Ка+ в жидкости, выбрасываемой из выделительной вакуоли, выше, чем в цитоплазме. Это показывает, что по крайней мере часть молекул N8 тАУ К-насоса размещена на мембране выделительной вакуоли. Значит, выделительная вакуоль регулирует не только содержание воды в инфузории, но и содержание Ш+. Таким образом, вакуоль гораздо больше похожа на почки высших организмов, чем думали раньше.
Посмотрим теперь, как Ка+ попадает внутрь клетки. Во-первых, он заглатывается вместе с пищей и попадает в пищеварительные вакуоли. В вакуоли поступают пищеварительные ферменты и идет переваривание пищи; в некоторый момент путешествия вакуоли там создается кислая среда, как в желудке человека, а потом среда в ней становится щелочной и таким образом Влпутешествующий желудокВ» превращается в Влдвенадцатиперстную кишкуВ». У человека пища подвергается разным воздействиям в разных точках пищеварительного тракта, а у инфузорий тАУ в разные моменты времени в одном и том же мембранном пузырьке. Но, несмотря на эти различия, результат один: пища в вакуоли расщепляется до аминокислот и других мелких молекул. В мембране вакуолей имеются разнообразные транспортные молекулы, ведь молекулы углеводов и аминокислот должны быть переправлены в цитоплазму до того* как пищеварительная вакуоль окончит свое существование и сольется с наружной мембраной. Среди этих транспортных молекул много Влэлектрических транспортеровВ», которые присоединяют к себе молекулы пищевых веществ и ион Ма+, а затем выбрасывают их в цитоплазму. Мы рассказывали вам о работе таких транспортеров в параграфе ВлЗачем невозбудимым клеткам потенциал покояВ». Разница состоит лишь в том, что там мы говорили о транспорте веществ из наружной среды в клетку, а у инфузорий этот транспорт идет из пищеварительной вакуоли. Но это различие несущественно, можно считать, что в таких вакуолях находится кусочек проглоченной внешней среды. Вот таким образом вместе с молекулами сахара и аминокислот ионы + и попадают в протоплазму. А дальше к ним вполне можно применить выражение: ВлНатрий сделал свое дело, натрий может уходитьВ». Только уходят они не сами, а выкачиваются натриевым насосом в выделительную вакуоль.
Существует и второй путь поступления] + в клетку инфузории. Мы уже говорили, что у инфузории есть разные Влорганы чувствВ». Расмотрим тут ее чувствительность к механическим воздействиям. При прикосновении к переднему концу инфузории в ней возникает деполяризационный рецепторный потенциал; а если Влудар по носуВ» достаточно силен, то возникает и ПД. Эти потенциалы создаются в основном ионами Са++, которых в пресной воде больше. Однако при деполяризации или ПД в протоплазму клетки попадают и ионы Лта+.
Проникновение Са++ внутрь клетки влияет на орган движения инфузории тАУ реснички тАУ точно так же, как у нас вхождение Са++ в мышечные клетки необходимо для их сокращения. Кстати, и разрядка трихоцист у инфузорий связана с потоками Са++ в клетку. Большинство Са-каналов инфузорий расположено прямо на мембране ресничек. Если удар по передней части инфузории вызвал ПД, то откроется много Са-каналов, внутрь клетки войдет много Са++, а от этого реснички инфузории меняют направление своего удара. Возникает ВлреверсВ»: инфузория отплывает Влхвостом впередВ» от раздражителя, например от препятствия, на которое она натолкнулась. После того как Са-насос и митохондрии уберут излишки Са++ из цитоплазмы, нормальная работа ресничек восстанавливается.
Изучение плавательного поведения туфельки показало, что работа ее ресничек зависит от МП. При нормальном
ГШ ее реснички ударяют примерно 20 раз в секунду, а при гиперполяризации частота их возрастает до 50 в секунду. При сильной деполяризации, как мы уже говорили, возникает ВлреверсВ». В действительности движением ресничек управляет не сам потенциал, а концентрация ионов Са++ и других веществ. Например, при гиперполяризации туфельки, как и при деполяризации, в клетку входят ионы Са++, но они входят через особые каналы, расположенные иначе, и это вызывает не реверс движения, а ускорение ударов ресничек. Реснички покрывают фактически все тело инфузории, поэтому их каналами удобно управлять с помощью МП; ведь туфелька тАУ одна небольшая клетка и изменение потенциала в одной ее части распространяется и на другие. Отсюда ясно, что управлять с помощью МП разнообразными процессами для туфельки невозможно: процессов, которыми надо управлять, много, а потенциал один. Туфелька выходит из положения темВ» что использует ионные каналы, чувствительные не к сдвигу МП, а другим сигналам, например, каналы трихоцист чувствительны к механическим или химическим воздействиям.
Было выяснено, что в ВлхвостеВ» туфельки имеется чувствительный участок, при механическом раздражении которого открываются К-каналы и возникает гиперполяризация клетки, что увеличивает частоту работы ресничек. В результате этого нормальная туфелька начинает плыть быстрее, если ее хватают за хвост.
Мы сказали слова Влнормальная туфелькаВ» не случайно. Дело в том, что существуют инфузории-мутанты. При их изучении было показано, что мутация в одном гене может сделать дефектным белок, образующий тот или иной канал. Существует мутация, которая портит Са-канал, чувствительный к удару по передней части тела инфузории. Такая инфузория, наткнувшись на препятствие, все время Влбьется головой о стенуВ», так как ионы Са++ в нее при механическом раздражении не входят и не обеспечивают реверс. Другая мутация портит Са-канал иначе, замедляя его инактивацию. Такие мутанты становятся очень ВлпугливыВ»: столкнувшись с препятствием, они отскакивают от него и долго-долго плывут хвостом вперед, так как через их Са-каналы входит много Са++ и нормальная работа ресничек долго не восстанавливается.
Интересно, что передний и задний концы туфельки по-разному реагируют не только на механические, но и на температурные воздействия. В 1987 г. японские исследователи показали, что на переднем конце туфельки имеется участок, чувствительный к холоду, а на заднем тАУ к теплу; у позвоночных животных для той же цели служат специальные клетки тАУ тепловые и холодовые рецепторы. При действии на туфельку холода на ее переднем конце открываются Са-каналы, а при действии тепла на заднем закрываются К-каналы. Результат один и тот же: МП снижается. Однако поведенческий эффект температурного воздействия состоит в основном не в изменении скорости плавания, а в том, что туфелька начинает заметно чаще, чем при привычной для нее температуре, менять направление своего движения, Если при таких метаниях она выскочит вновь в область привычных температур, то поплывет по прямой, удаляясь от горячего или холодного места. Как управляется частота поворотов, пока неизвестно. Замечательная особенность ионных каналов, чувствительных к температуре, состоит в том, что они обладают памятью. Если инфузория жила при 20 С, то 15 С для нее холодно, и реагируют холодовые рецепторы; но если инфузория достаточно долго жила при 10 С, то 15 С для нее тепло, и реагируют тепловые рецепторы. Как перестраивается работа чувствительных к температуре каналов во время привыкания, пока не выяснено.
У такая мозаика особенно богата, так как она должна обеспечить этой клетке разнообразное поведение. Так что инфузория тАУ интереснейший пример Влмозаики каналовВ». С другой стороны, изучение инфузорий дает нам еще один урок. Совместная работа ионных каналов туфельки весьма затрудняет их изучение. Мы так много поняли о туфельке потому, что до этого были изучены клетки других разнообразных животных. Очень полезно сравнивать разные объекты. Оказалось, что каналы и органеллы туфельки работают в общем на тех же принципах, что и каналы и органы других организмов. Очень интересно было бы выяснить, какие черты этого сходства объясняются наследством, полученным от общего предка, а какие являются результатом естественного отбора, который привел по разным путям к сходным результатам,
Об электростанциях клеток и тАУ немного о бактериях тАУ первых электриках Земли
Без этого небольшого параграфа наша книга была бы неполна, ведь в ней отсутствовало бы важнейшее для всего живого использование электричества тАУ использование его для обеспечения каждой клетки энергией.
Откуда организмы получают энергию? Животные тАУ из пищи, которую они съедают. Биохимиками было выяснено, что пища у животных, дышащих кислородом воздуха, медленно окисляется, а за счет этой энергии в организме синтезируется особое вещество тАУ АТФ. Это вещество играет роль универсальной валюты и расходуется животными для синтеза новых веществ, для работы мышц или ионных насосов и т.д. Было выяснено также, что процесс сгорания пищи и синтеза АТФ осуществляется особыми клеточными органеллами тАУ митохондриями, описанными еще в 1850 г. Келликером в мышцах насекомых. Митохондрии имеются не только у животных, но и у растений и грибов, нет их только у бактерий.
Если сделать еще один шаг в своем любопытстве и задать вопрос: ВлА откуда же берется пища?В», то на него ответ тоже известен. В конечном счете пища производится фотосинтезирующими организмами, получающими свою энергию от Солнца. В растениях имеются, кроме митохондрий, особые органеллы тАУ хлоропласта, содержащие хлорофилл. Эти органеллы умеют, как и митохондрии, синтезировать АТФ, а кроме того, умеют синтезировать из воды и углекислого газа углеводы.
Все реакции окисления пищи, синтеза АТФ, синтеза углеводов достаточно сложны, происходят при участии многочисленных ферментов и более полувека широко изучаются в разнообразных биохимических лабораториях. Какое же отношение они имеют к теме этой книги? Еще лет 30 назад на этот вопрос был бы дан уверенный ответ тАУ никакого! Но в 1961 г. английский ученый Ж. Митчел выдвинул гипотезу, что энергия пищи сначала преобразуется в электрическую энергию, а уж та затрачивается на производство АТФ. Эта гипотеза была в конце концов доказана, и мы считаем необходимым кратко рассказать об энергетическом обеспечении организмов. А кратким наш рассказ будет прежде всего потому, что на эту тему недавно была написана обстоятельная, понятная и яркая книга ВлРассказы о биоэнергетикеВ», автор которой В.П. Скулачев тАУ один из создателей биоэнергетики.
В чем же более конкретно состояла гипотеза П. Митчела? Митчел предположил, что окисление пищи приводит к возникновению разности потенциалов на мембране митохондрий за счет выхода протонов из митохондрий, затем эта электрическая энергия тратится
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)