Что такое ген? Генетическая точка зрения
РЕФЕРАТ
ЧТО ТАКОЕ ГЕН? ГЕНЕТИЧЕСКАЯ ТОЧКА ЗРЕНИЯ
Введение
В концепции гена сфокусированы результаты примерно ста лет работы, потребовавшейся для того, чтобы разгадать основу наследственности. Проведенные исследования были столь многоплановы, что нельзя однозначно и удовлетворительно ответить на вопрос: что такое ген?
Ген - это последовательность ДНК, несущая информацию об определенном белке. Еще совсем недавно это казалось вполне достаточным (если не полным) биохимическим определением. Последовательность ДНК можно было бы определить как непрерывную линейную последовательность нуклеотидов, колинеарную аминокислотной последовательности соответствующих белков. Однако, как теперь уже установлено, последовательность, в которой закодирован белок, не всегда непрерывна. Она может прерываться вкрапленными в нее некодирующими участками. Таким образом, гены могут состоять из отдельных кусков, соединяющихся воедино в процессе генной экспрессии.
Ген можно идентифицировать по кластеру (группе) мутаций, каждая из которых предотвращает образование соответствующего белка. Хотя данное положение остается справедливым и в отношении прерывистых генов, свойства их кластеров в этом случае оказываются значительно сложнее. Такое утверждение достаточно спорно, поскольку большинство прерывистых генов обнаружено в таких ситуациях, в которых детальный генетический анализ невозможен. Однако очевидно, что генетический взгляд на ген необходимо пересмотреть.
У всех видов большое число генов, составляющих геном, организовано в сравнительно небольшое число хромосом. Генетический материал каждой хромосомы представляет собой чрезвычайно длинную последовательность ДНК, содержащую множество линейно расположенных генов. Довольно долго оставалось загадкой, сколько всего генов существует в геноме. Представление о том, что каждый ген может существовать сам по себе, как уникальное целое, пришлось пересмотреть, когда было обнаружено, что во многих случаях кластеры родственных генов организованы в небольшие семейства.
Обычно считалось, что геном довольно стабилен и изменения в его общей структуре и организации происходят крайне редко - лишь в растянутой эволюционно-временной шкале. Однако это противоречит недавно полученным данным о том, что в некоторых случаях в ДНК периодически происходят перестройки и что существуют относительно мобильные компоненты генома.
Сама концепция гена, таким образом, подверглась эволюции. И хотя многие из традиционных положений данной концепции остались без изменений, были обнаружены исключения, свидетельствующие о том, что ни одно из положений нельзя считать абсолютным. Со времени открытия гена как неизменной единицы наследственности его свойства описывали, исходя из того, что положение гена в хромосоме фиксировано. Из этого в свою очередь было развито представление о том, что генетический материал хромосомы представлен непрерывной нитью ДНК, соответствующей множеству генов. Следуя за развитием этих идей, мы сможем дать операциональное описание гена, но это вряд ли поможет нам сформулировать краткое определение.
Элементарный фактор наследственности
"Постоянные признаки, появляющиеся в данной группе растений, могут быть получены во всех комбинациях в соответствии с математическими законами сочетаний. Те признаки, которые при передаче потомству проявляются и, следовательно, обнаруживаются у гибридов, называют доминантными, а те, которые оказываются в скрытом состоянии, называют рецессивными. Выражение "рецессивный" было выбрано потому, что такие признаки как бы отступают или полностью исчезают у гибридов, но вновь появляются в их потомстве без каких-либо существенных изменений и в предсказуемом соотношении. Промежуточные формы ни в одном из экспериментов обнаружены не были."
Эта (сокращенная) цитата из замечательной статьи Менделя, опубликованной в 1865 г., вводит основную концепцию генетики: существует единица наследственности в виде некоторого фактора, который передается от родителей потомству. Этот дискретный фактор не что иное, как хорошо нам теперь известный ген. Работа Менделя пролила свет на общее поведение гена при наследовании в ряду поколений. Из этой работы следовало, что отдельный живой организм-это способ, которым ген экспрессируется и увековечивает себя. Суть этого представления изображена на рис.1 в виде чередования поколений.
В отличие от запутанных ранее представлений о наследственности работа Менделя утвердила генетику как экспериментальную науку, введя в нее несколько новых подходов. Во-первых, исходный набор растений был "чистым по признаку": каждое растение давало в потомстве только растения с данным признаком и никогда-с противоположным. Сначала было изучено наследование только одного признака. После этого можно было одновременно исследовать наследование двух признаков или более. Свои эксперименты Мендель поставил на количественную основу, проводя учет потомков каждого типа в каждом скрещивании.
Свойства генов, выявленные в работе Менделя, сформулированы в его первом и втором законах. Первый закон рассматривает особенности индивидуального гена. Организм имеет две копии каждого гена, т.е., выражаясь современным языком, он диплоиден. Только одна из двух копий гена попадает от родителя к потомку через гаметы (половые клетки). Гаметы, сливаясь, образуют зиготу (оплодотворенное яйцо), несущую по одной копии от каждого родителя. В результате каждый организм получит одну отцовскую копию гена и одну материнскую.
Ген может существовать в альтернативных формах, что проявляется в различии признаков - например, красная окраска цветка в противовес белой.
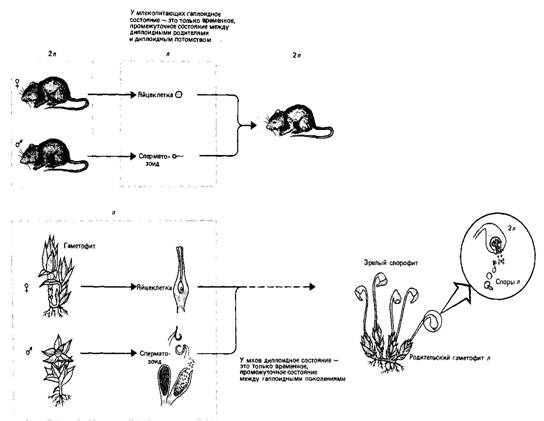
Рис. 1. Когда эукариоты "увековечивают" свои гены путем чередования диплоидного и гаплоидного состояний, одно из состояний преобладает у "взрослого" организма, тогда как другое связано с образованием гамет или зигот. Млекопитающие и мхи - это крайние примеры. У первых взрослое поколение диплоидно, у вторых - гаплоидно.
Красным цветом указаны диплоидные (2и) ткани, серым - гаплоидные (и) называют аллелями. Закон независимого расщепления утверждает, что присутствуя одновременно в одном растении, эти аллели не изменяют друг друга, но расщепляются, переходя в различные гаметы, при формировании следующей генерации.
У организма, чистого по признаку, т.е. у гомозиготы оба аллеля одинаковы. Но скрещивание двух родителей, каждый из которых гомозиготен по разным аллелям, приведет к образованию гибрида, или гетерозиготы. Если один аллель доминантен, а другой рецессивен, то организм по своему внешнему виду, или фенотипу, будет только доминантного типа (так что гетерозиготы не отличаются от чистого по признаку доминантного родителя).
Но, согласно первому закону Менделя, генетическая конституция, или генотип, гибрида включает в себя оба аллеля. Как показано на рис.2, это можно установить, если скрестить между собой два гибрида и получить второе поколение. Критическое значение при этом имеет тот факт, что аллели не смешиваются друг с другом ("промежуточных форм не обнаружено"), а сохраняют свою физическую целостность, взаимодействуя только на уровне экспрессии.
Хотя в опытах Менделя исследованные признаки проявляли (к счастью) полное доминирование, так бывает не всегда. Доминирование аллелей может быть неполным (частичным) или совсем отсутствовать (что иногда называют ко доминантностью). В последнем случае гетерозиготы отличаются от гомозигот промежуточными свойствами. Например, скрещивание растений львиного зева с красными и белыми цветами дает розовые гибриды. Хотя в этом случае нет доминирования, наблюдается все та же закономерность - гибриды первого поколения (F1) одинаковы. В дальнейших скрещиваниях гибридов образуются те же численные соотношения, как и при полном доминировании, хотя вместо л чух фенотипов можно различить три.
Независимость различных генов
Во втором законе Менделя утверждается независимость комбинации различных генов. При скрещивании растения, доминантного по двум различным признакам, с растением, у которого эти признаки рецессивны, F1 по-прежнему состоит из одинаковых растений доминантного типа. Но при последующем скрещивании гибридов F1 появляются два класса растений. Один класс состоит из растений исходных родительских типов. Второй класс включает растения новых фенотипов, с доминантными признаками одного родителя и рецессивными признаками другого. Эти фенотипы называют рекомбинантными.
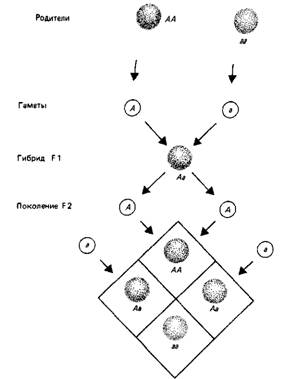
Рис.2. Аллели расщепляются независимо.
Два родителя гомозиготны: А А имеет две копии доминантного аллеля; аа имеет две копии рецессивного аллеля. Каждый образует только один тип гамет, так что F1 (первое гибридное поколение) состоит из одинаковых гибридных особей Аа. Поскольку А доминирует над а, фенотип потомков Аа такой же, как у родителя АА (показано цветом). Фенотип рецессивной гомозиготы аа показан отсутствием цвета. Каждый гибрид F1 образует гаметы А и а в одинаковом количестве. При скрещивании объединение этих гамет приведет к образованию F2 (второе гибридное поколение), которое будет состоять из \АА: 2Аа: \аа. Так как АА и Аа имеют одинаковый фенотип, получается классическое соотношение доминантного и рецессивного фенотипов-3:
1. В тех случаях, когда фенотип гетерозиготы занимает промежуточное положение между родительскими АА и аа, F1 будет другим, а в F2 отношение фенотипов будет: 1 доминантный: 2 промежуточных:! рецессивный.
На рис.3 показано, что соотношение потомков различного типа можно объяснить, предположив, что в процессе образования гамет происходит совершенно случайное объединение одного из аллелей первого признака с одним из аллелей второго. Четыре возможных типа гамет образуются в равных соотношениях; при формировании зиготы следующего поколения они объединяются случайно.
Типичное соотношение фенотипов скрывает за собой большее разнообразие генотипов, что может быть установлено путем возвратного (анализирующего) скрещивания с рецессивным родителем. По существу, возвратное скрещивание позволяет прямо установить генотип исследуемого организма (рис.4).
Закон независимого распределения генов, как следует из сказанного выше, говорит, что поведение любой пары (или большего числа) генов можно полностью предсказать с помощью правил комбинаторики. Распределение между потомками (наследование) одного гена не влияет на распределение другого. Эта концепция подразумевает, что распределение генов происходит со статистической вероятностью, а не предопределено точно. Чем больше проведено скрещиваний, тем точнее будет совпадать с предсказанным соотношение типов в потомстве.
Роль хромосом в наследственности
Открытие Менделя не сразу оценили по достоинству, поскольку не было никаких представлений о физической основе постулируемых факторов. Хромосомная теория наследственности, предложенная одновременно Саттоном и Бовери (Sutton, Boveri), разрешила долгие споры о возможной роли хромосом. Уже существовали некоторые, хотя довольно туманные, предположения о том, что хромосомы как-то участвуют в механизмах наследования. В 1903 г. наконец стало ясно, что по своим свойствам они точно совпадают с дискретными единицами наследственности, описанными Менделем.
Клеточная теория, утвердившаяся в середине девятнадцатого века, предполагала, что все организмы состоят из клеток и что эти клетки могут возникать только из предсуществующих клеток. Ранние цитологические исследования показали, что "типичная" клетка состоит из плотного ядра, отделенного мембраной, от менее плотной окружающей цитоплазмы. Внутри ядра с помощью определенных красителей можно было различить зернистые участки - хроматин. Вскоре после работы Менделя было установлено, что хроматин представлен определенным числом нитевидных образований, или хромосом.
В большинстве клеток хромосомы видны только в процессе клеточного деления. Два типа деления, присущие организмам, размножающимся половым путем, объясняют и постоянство генетического материала, и процесс наследования в том виде, как это было предсказано законами Менделя.
Клеточный цикл у растущего организма состоит из двух этапов. Более длинный период-интерфаза, когда клетка синтетически активна и занята воспроизведением своих компонентов. Затем следует короткий период-митоз, интерлюдия, во время которой фактически завершается процесс разделения на две дочерние клетки. Клетки, возникающие в результате ряда митотических делений и образующие целый организм, называют соматическими клетками.
Каждая дочерняя клетка, начинающая свою жизнь после митоза, содержит по две копии каждой хромосомы. Их называют гомологичными. Общее число хромосом в клетке, известное как диплоидный набор, обозначают 2п. Типичная соматическая клетка существует в диплоидном состоянии (кроме того периода, когда она готовится к делению или уже делится).
В интерфазе растущая клетка удваивает свой хромосомный материал. Однако это становится очевидным только в последующем митозе. В митозе каждая хромосома разделяется вдоль по длине, образуя две копии - сестринские хроматиды. В этот момент клетка содержит 4и хромосом, организованных в 2и пар сестринских хроматид. Иными словами, в клетке имеется по две (гомологичные) копии каждой пары сестринских хроматид. На рис.5 показана последовательность процессов, обеспечивающих митотическое деление. Суть заключается в том, что сестринские хроматиды растаскиваются к противоположным полюсам клетки, так что каждая дочерняя клетка получает по одной копии каждой сестринской хроматиды. Теперь это самостоятельные хромосомы.4и хромосомы, существовавшие в начале деления, разделились на два набора по 2и хромосом. Этот процесс повторяется в следующем клеточном цикле. Таким образом, митотическое деление гарантирует постоянство набора хромосом в соматических клетках.
Один родитель гомозиготен по двум доминантным генам - по гену А, определяющему цвет, и гену В, определяющему форму (круглая форма). Другой родитель гомозиготен по рецессивным аллелям а и Ъ (отсутствие цвета, морщинистая форма). Потомство F1 одинаково-с доминантным фенотипом.
Родители F1 образуют гаметы, в которых происходят независимое расщепление аллелей и независимая комбинация генов, так что образуется равное количество гамет каждого из четырех возможных типов. Они соединяются случайно и образуют 9 генотипических классов, которые из-за отношений доминантности проявляются в виде четырех фенотипических классов: 9 окрашенных гладких: 3 окрашенных морщинистых: 3 бесцветных гладких: 1 бесцветный морщинистый. Заметим, что каждый реципрокный генотип представлен одинаковым числом особей, например два родительского типа (по одному ААВВ и ааЪЬ) и два рекомбинантных (по одному каждого рекомбинантного класса ААЪЬ и ааВВ). Соотношение 3: 1 сохраняется для каждого независимо расщепляющегося признака.
Число фенотипических классов будет больше, если один или оба признака не проявляют полного доминирования (в этом случае гетерозиготы отличаются от любой гомозиготы). Их будет меньше, если два гена действуют на один признак. Так, если бы оба гена А а В были нужны для развития окраски, то у гибридов F2 наблюдалось бы соотношение 9 окрашенных: 7 бесцветных.
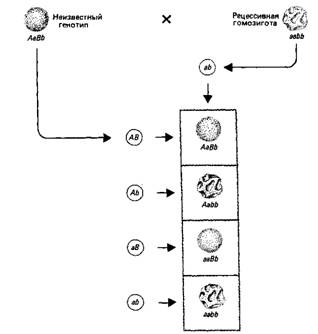
Рис. 3.
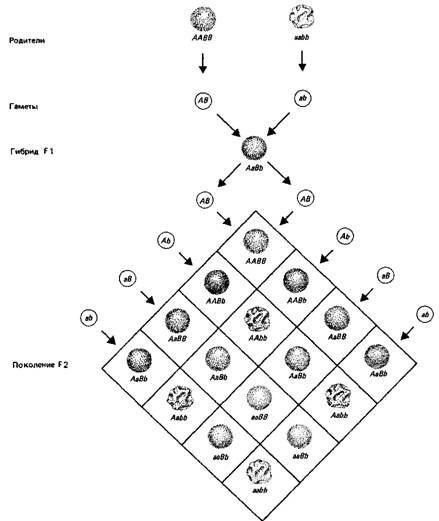
Рис.4. Возвратное скрещивание с рецессивной гомозиготой используется для определения генотипа.
Соотношение гамет, образованных растением с неизвестным генотипом, отражает обычно независимое расщепление аллелей и независимую комбинацию генов. Генотип каждой гаметы выясняется непосредственно в скрещивании с гаметой, несущей только рецессивные аллели.
Иной цели служит процесс образования половых клеток (гамет, т.е. яйцеклеток или сперматозоидов), который происходит при другом типе деления. Этот путь деления изображен на Рис.6. В процессе мейоза образуются клетки, содержащие гаплоидное число (и) хромосом. Этот процесс включает в себя два последовательных деления. И в этом случае хромосомы сначала удваиваются, так что клетка приступает к делению в состоянии 4и.
В начале первого деления гомологичные пары сестринских хроматид конъюгируют (соединяются попарно), образуя биваленты. Каждый бивалент содержит все четыре копии одного гомолога, находящиеся в клетке. В первом делении каждый бивалент расщепляется на две пары сестринских хроматид, разделенных в свою очередь на полухроматиды. В результате образуются два набора по 2п хромосом, из з пар сестринских хроматид каждый.
Далее следует второе мейотическое деление, в котором оба набора 2п делятся еще раз. Это деление формально подобно митотическому делению, поскольку в различные дочерние клетки попадает по одной из копий каждой пары сестринских хромосом.
Общий результат мейоза заключается в том, что исходное число хромосом 4п делится на четыре гаплоидные клетки, дающие начало зрелым яйцеклеткам или сперматозоидам.
При формировании гамет гомологи отцовского и материнского происхождения разделяются так, что каждая гамета получает только один из двух гомологов своих родителей. Существенным моментом в этом процессе является тот факт, что в полном соответствии с законами Менделя наблюдается независимость сегрегации негомологичных хромосом. Каждый член одной гомологичной пары входит в гамету в случайном сочетании с любым членом другой гомологичной пары.
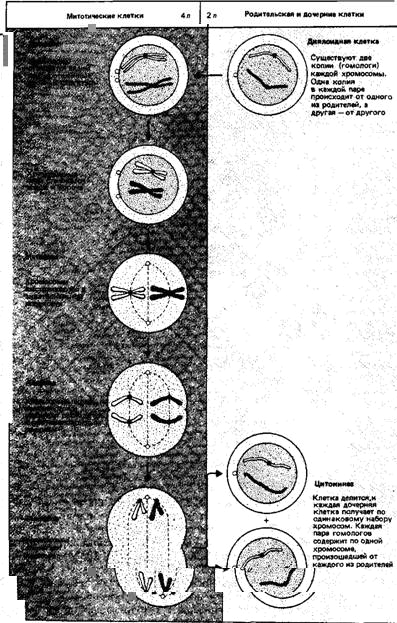
Рис. 5. Митоз обеспечивает постоянство хромосомного состава клетки. Одна пара гомологичных хромосом (обозначена белым) происходит от одного из родителей, другая (окрашена) - от второго родителя. Диплоидная клетка имеет по две копии каждой хромосомы (общее число 2п). Перед митозом хромосомы удваиваются, и профаза, следовательно, начинается с состояния 4п. Два члена каждой пары разделяются на две отдельные хромосомы, которые затем расходятся в разные дочерние клетки. При этом восстанавливается диплоидное число 2п, так что дочерние клетки имеют тот же набор хромосом, что и родительская клетка.
Подытожим параллели между хромосомами и менделеевскими единицами наследования. Гены встречаются в аллельных парах - по одному аллелю от каждого родителя в каждой паре; диплоидный набор хромосом образуется из двух гаплоидных родительских наборов. Распределение неаллельных генов в гаметах происходит независимо; негомологичные хромосомы подвергаются независимой сегрегации. Критическое условие, заключающееся в том, чтобы каждая гамета получала полный гаплоидный набор, выполняется независимо от того, рассматриваем ли мы этот процесс как сегрегацию хромосом или как распределение элементарных факторов наследственности.
Гены располагаются в хромосомах
Только установив, что определенный ген всегда присутствует в определенной хромосоме, можно доказать, что гены находятся в хромосомах. Такое доказательство было получено благодаря свойствам одного варианта плодовой мушки Drosophila melanogaster, обнаруженного Морганом в 1910 г. Этот белоглазый самец появился спонтанно в линии мух с обычными красными глазами.
Поскольку обычно встречаются мухи с красными глазами, их назвали диким типом; белые же глаза - это мутантный фенотип. Событие, приводящее к возникновению мутантного фенотипа, представляет собой генетическое изменение,, называемое мутацией. В результате большинства мутаций (но не всех) функционирование соответствующего гена нарушается частично или полностью. Поэтому изучение мутаций, как правило, сводится к изучению неактивных аллелей. Иногда происходят обратные мутации (реверсии), т.е. такие изменения в генетическом материале, которые восстанавливают исходное состояние, - явление, называемое реверсией к дикому типу.
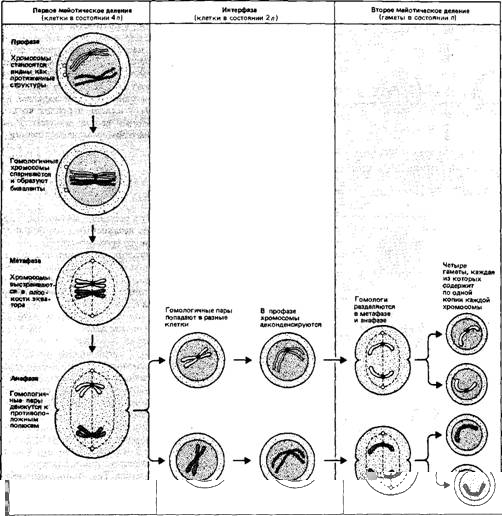
Рис. 6. В мейозе число хромосом уменьшается вдвое. На рисунке показано поведение одной пары гомологичных хромосом. Оба члена пары были удвоены перед началом профазы. Во время первого деления происходит спаривание гомологичных хромосом, а затем каждая пара расходится в разные клетки. Во время второго деления дуплицированные члены каждой пары расходятся в разные клетки, так что каждая гамета получает по одной копии. В клетке обычно находится несколько пар гомологичных хромосом, и члены каждой пары комбинируются между собой независимо друг от друга. Таким образом, подбор гомологичных хромосом (материнской и отцовской) в каждой гамете происходит случайно.
Мутацию white удалось локализовать в определенной хромосоме потому, что она оказалась связанной с полом. Обычно гомологичные пары хромосом при расхождении в мейозе образуют идентичные гаплоидные наборы. Однако у многих организмов, размножающихся половым путем, существует одно исключение из этого общего правила: самцы и самки могут иметь видимые различия в хромосомных наборах. Чаще всего у одного из полов одна хромосомная пара состоит из разных хромосом.
Такую пару называют половыми хромосомами, а остальные гомологичные пары - аутосомами. Хромосомные наборы двух полов можно записать следующим образом: 2А + XX и 2А + XY, где гаплоидный набор аутосом обозначают как А, а две половые хромосомы - как X и Х Пол с хромосомным набором 2А + XX называют гомогаметным; у него образуются только гаметы А + Х. Пол с набором 2А + Ч Х называют гетерогаметным, так как у него образуются в одинаковом количестве гаметы двух типов: А + Х и Б+Х. В результате случайного со единения гамет одного пола с гаметами другого пола постоянно сохраняется равное соотношение полов при образовании зигот. У Drosophila гомогаметным полом являются самки.
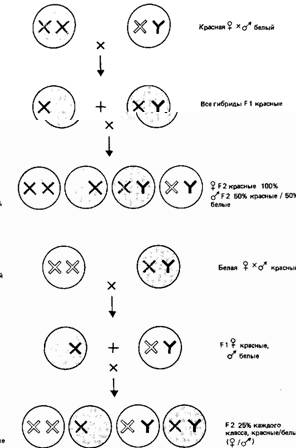
Хромосомы с красным аллелем (доминантным)
Хромосома с белым аллелем (рецессивным) Х Отсутствие аллеля в хромосоме (= рецессивному аллелю) О Красноглазая муха СЗ Белоглазая муха
Рис.7. Гены Х-хромосомы наследуются сцеплено с полом. Красная или белая окраска глаз у самца зависит только от Х-хромосомы, полученной им от матери. Фенотип самки зависит от того, получит ли она доминантный аллель от одного из родителей. При этом наблюдается характерный тип перекрестного наследования, сцепленного с полом, - от отца к дочерям, от матери к сыновьям (крисс-кросс).
Из законов Менделя следует, что результаты генетического скрещивания должны быть одинаковы независимо от того, кому из родителей принадлежит данный аллель. Но реципрокные скрещивания с признаком "белые глаза" у Drosophila дают различные результаты (рис.7).
Скрещивание белоглазый самец ч красноглазая самка дает лишь красноглазое потомство F1, что и можно было предсказать, если красный цвет доминантен, а белый рецессивен. Но в потомстве F2 все появившиеся белоглазые мухи оказались самцами. В реципрокном скрещивании красноглазые самцы ч белоглазые самки все самцы F1 были с белыми глазами, а все самки-с красными. При их скрещивании в потомстве F2 в равном соотношении появились белые и красные глаза у обоих полов.
Этот пример наследования строго обусловлен половыми хромосомами. Если аллели красных и белых глаз находятся в Х-хромосоме, и при этом Х-хромосома вообще не содержит локуса для окраски глаз, фенотип самца будет определяться единственным аллелем, находящимся в Х-хромосоме. Этот аллель перейдет ко всем дочерям, но ни один из сыновей его не получит. Данный случай является типичным примером наследования, сцепленного с полом.
Гены линейно выстроены вдоль хромосом
Независимая сегрегация хромосом, происходящая в мейозе, объясняет независимую комбинацию генов, расположенных в разных хромосомах. Но, как известно, общее число генетических факторов значительно больше, чем число хромосом. Следовательно, если все гены расположены в хромосомах, то каждая хромосома должна содержать много генов. Каковы же взаимоотношения между этими генами?
О стремительном темпе развития генетики в начале века свидетельствует тот факт, что в 1913 г., всего лишь через три года после сообщения о первом белоглазом мутанте, Стертевант (Sturtevant) сообщил об изучении наследования шести сцепленных с полом мутаций. В 1911 г. Морган (Morgan) показал, что каждый из этих факторов ведет себя в скрещиваниях так же, как красная и белая окраска глаз. Следовательно, каждый фактор должен располагаться в Х-хромосоме.
Некоторые из факторов оказались связанными между собой. Вопреки второму закону Менделя, доля родительских генотипов в F2 оказалась выше предполагаемой, что объяснялось пониженным образованием рекомбинантных типов в этом поколении. Тенденция некоторых признаков оставаться связанными, вместо того чтобы комбинироваться независимо, была названа сцеплением. На рис.8 показано, как измеряют сцепление.
Морган предположил, что причина генетического сцепления факторов - это "просто механический результат их локализации в одной хромосоме". Он предположил также, что образование генетических рекомбинантов можно отождествить с процессом кроссинговера, наблюдаемого в мейозе. В раннем мейозе, на стадии, когда четыре копии каждой хромосомы представлены в виде бивалента, между близко расположенными (конъюгировавшими) гомологичными парами происходит попарный перекрест генетического материала, названный хиазмой. Этот процесс схематически изображен на рис.9.
Забежим на несколько лет вперед. Только в 1931 г. было формально доказано, что рекомбинация обусловлена кроссинговером. У кукурузы и у дрозофилы были получены подходящие транслокации, при которых оторванная часть одной хромосомы прикрепилась к другой. Это позволяет отличить по внешнему виду хромосому с транслокацией от нормальной хромосомы. Как изображено на рис.10, в подобных скрещиваниях можно показать, что образование генетических рекомбинантов происходит только при физическом обмене между соответствующими областями хромосом.
Хромосомы изображены в виде окрашенных полосок. Гены гетерозиготного родителя обозначены черными буквами, гены родителя, рецессивного по двум признакам, цветными буквами. Это скрещивание относится к тому же типу, что и скрещивание, показанное на рис.4. В скрещивании, изображенном сверху, одна хромосома несет оба доминантных аллеля, другая - оба рецессивных (скрещивание с аллелями "в фазе притяжения"). Таким образом, родительские типы представляют собой АВ и ab, рекомбинантные типы-zli и аВ.
В скрещивании, изображенном снизу, одна хромосома гетерозиготы несет один доминантный и один рецессивный аллель, а другая хромосома несет противоположную комбинацию (скрещивание с аллелями "в фазе отталкивания"). Следовательно, родительские типы - это АЬ и аВ, рекомбинантные - АВ и аЪ. В каждом случае в потомстве наблюдается увеличение доли родительских типов (70%) и уменьшение доли рекомбинантных типов (30%) по сравнению с 50% для каждого типа, что ожидалось бы при независимой комбинации генов. Заметим, что в потомстве, полученном от этих скрещиваний, два родительских типа присутствуют в одинаковом количестве; одинаково и количество двух рекомбинантных типов. Сцепление в данном случае между А и В равно 30%, или 30 условным единицам карты.
Если вероятность образования хиазмы между двумя точками в хромосоме зависит от расстояния между ними, гены, расположенные ближе друг к другу, будут наследоваться вместе. По мере увеличения расстояния между двумя генами возрастает и вероятность кроссинговера между ними. Таким образом, если рекомбинация обусловлена кроссинговером, гены, находящиеся близко друг к другу, должны быть тесно сцеплены, причем генетическое сцепление будет уменьшаться по мере физического удаления. И наоборот, генетическое сцепление можно использовать как меру физического расстояния.
Исходя из представления о том, что гены одной хромосомы сцеплены между собой, Стертевант (Sturtevant) предложил использовать частоту рекомбинации в качестве единицы расстояния на карте для измерения относительной локализации генов. Эта единица расстояния выражается как процент рекомбинации:
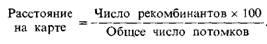
Расстояние измеряется в условных единицах карты;одна единица карты (сантиморганида) соответствует 1% рекомбинантных потомков. Основные закономерности рекомбинации были установлены при изучении всех шести сцепленных с полом признаков вместе. Индивидуальные расстояния на карте аддитивны. Иными словами, если два гена А и В находятся на расстоянии 10 ед. друг от друга, а расстояние от в до С равно 5 ед., то расстояние между А и С, полученное при прямом измерении, будет близко к 15. Поэтому гены можно расположить в линейном порядке.
Критическое значение для построения генных карт имеет тот факт, что расстояние между генами не зависит от использованных аллелей, а зависит только от положения генных локусов. Локус-это место в хромосоме, в котором находится ген данного признака. Все различные альтернативные формы гена, т.е. аллели, используемые при картировании, находятся в одном и том же локусе. Таким образом, генетическое картирование позволяет установить взаимную локализацию генных локусов, расположенных в хромосоме в определенном месте и в линейном порядке. В экспериментах по картированию один и тот же результат получается независимо от конкретной комбинации аллелей (рис.8). Стертевант заключил, что его результаты "служат новым доводом в пользу хромосомной теории наследственности, поскольку они убедительно показывают, по крайней мере математически, что исследуемые факторы расположены в виде линейных рядов".
Этот вывод представляет большой интерес. Можно построить генетическую карту, в которой хромосома будет изображена в виде линейного ряда генов. Однако это еще не доказывает, что генетическое содержание хромосомы представлено физически непрерывным рядом генов. Рассматривалось множество других моделей хромосомы, пока не стало очевидным, что хромосома состоит только из одной-единственной нити генетического материала.
Группа сцепления включает в себя все гены, которые прямо или опосредованно сцеплены между собой. Гены, расположенные рядом, сцеплены между собой прямо; гены же, отстоящие друг от друга более чем на 50 ед. карты, распределяются практически независимо. Поскольку сцепление действует на протяженном участке, каждый новый идентифицированный ген в организме можно поместить рядом с группой генов, уже картированных ранее. Таким образом, гены распадаются на дискретное число групп сцепления. Гены одной из групп сцепления всегда независимо комбинируются с генами других групп.
Число групп сцепления совпадает с числом хромосом. Относительная длина групп сцепления аналогична относительным размерам хромосом. На рис.11 в качестве примера рассматриваются хромосомы дрозофилы, у которых оказалось легко измерить длину. Сформулированная Менделем концепция гена как дискретного элементарного фактора наследственности может быть расширена в концепцию, рассматривающую хромосому как протяженную единицу, состоящую из многих генов. Физическое положение генов лежит в основе их генетического поведения.
Генетические карты непрерывны
На тех генетических картах высших организмов, которые были установлены в первой половине этого века, гены были расположены как бусы на нитке. Они были закреплены в определенном порядке, и генетическую рекомбинацию представляли как обмен соответствующими участками нитки между гомологичными хромосомами. При этом ген со всеми своими свойствами выступает как таинственный объект (бусинка), чье взаимоотношение с ее окружением (ниткой) совершенно не ясно.
Из-за небольшого числа потомков, получаемых в результате скрещивания, при построении рекомбинационных карт достигается лишь низкое разрешение.
Строение индивидуального локуса было исследовано в серии опытов с генами rll фага Т4, участвующими в лизисе бактериальной клетки. Мутации в этих генах вызывают быстрый лизис. Были подобраны такие условия опыта, когда при совместном инфицировании бактерий двумя гП-мутантами фага потомство образуется лишь при условии, что в результате рекомбинации мутантных фагов появится фаг дикого типа. Частота рекомбинации при этом также зависела от расстояния между мутациями, как и в случае эукариотической хромосомы.
Высокая разрешающая способность этого подхода, благодаря селективной методике, позволяет количественно оценить даже самые редкие рекомбинационные события, а это дает возможность измерять расстояние по карте для любой пары мутаций. (Единственным ограничением этого метода является частота обратных мутаций, или "реверсий", к дикому типу. Эта частота составляет примерно 0,0001%. Таким образом, чтобы достичь минимального уровня измерений, превышающего в 10 раз фон спонтанных мутаций, можно измерять только рекомбинационные частоты выше 0,001%)
Было проанализировано примерно 2400 мутаций, что дало возможность идентифицировать 304 различных мутантных сайта. (Если две мутации не рекомбинировали между собой, их принимали за две спонтанные мутации одного и того же генетического сайта) Исследованные мутации можно было расположить в линейном порядке. Из этого следовало, что сам ген имеет тот же линейный принцип устройства, что и гены в хромосоме. Из этих данных вытекает, что генетическая карта линейна как внутри генов, так и между ними, т.е. она представляет собой непрерывную последовательность, включающую в себя все гены.
Один ген - один белок
До 1945 г. было лишь известно, что гены - это основные единицы наследственности. Но каким путем они выполняют свою функцию, оставалось неясным. Гены можно было идентифицировать, только исходя из мутаций, вызывающих некоторые отклонения от нормы в фенотипе, причем степень отклонения варьировала от изменения одного признака (такого, как цвет глаз) до крайне резких морфологических перестроек, затрагивающих ряд тканей организма. Почти во всех случаях эффект мутации анализировали только на описательном уровне. Однако предполагали, что существует некоторая общая основа мутационных воздействий.
Идея о том, что действие мутационных факторов опосредовано ферментами, возникла почти одновременно с возрождением генетики. В работах, проведенных с 1902 по 1908 г., Геррод (Garrod) высказал мнение, что болезнь человека тАУ алкаптонурия - обусловлена нарушением какой-то метаболической реакции, катализируемой ферментом. Его фраза - "врожденные ошибки метаболизма" - заключает в себе концепцию, согласно которой генетический дефект может привести к нарушению определенного метаболического процесса, обусловливая тем самым наблюдаемый фенотип. В последующие три десятилетия накопились примеры влияния специфических мутаций на определенные биохимические реакции. Основная трудность исследований этого периода состояла в том, что приходилось довольствоваться случайно отобранными мутациями, не всегда пригодными для биохимического изучения.
С помощью рентгеновского облучения были индуцированы мутанты Neurospora, не способные расти на минимальной среде. Биохимическую природу дефекта можно было установить. Для этого было достаточно определить, добавление какого именно вещества в среду позволяет расти мутантному штамму. У каждого мутанта блокирована определенная метаболическая стадия и каждой такой стадии у штамма дикого типа соответствовал один определенный фермент. Если какой-то субстрат утилизируется в результате целой серии метаболических реакций, мутация, нарушающая любую из этих реакций, может блокировать всю серию реакций. Об этом свидетельствует накопление метаболического предшественника того соединения, которое образуется на блокированной ступени синтеза. На рис.12 изображен пример пути биосинтеза глазного пигмента дрозофилы, последовательные этапы которого хорошо согласуются с рассмотренной выше схемой.
К 1945 г. результаты этого анализа легли в основу гипотезы: "Один генодин фермент". Согласно этой гипотезе, каждую метаболическую ступень катализирует отдельный фермент, за образование которого отв
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)