Эволюция генома
Геномика - наука, которая изучает структурно-функциональную организацию генома, представляющего собой совокупность генов и генетических элементов, определяющих все признаки организма. Термин геномика появился в 1987 году, а первый полный бактериальный геном был расшифрован в 1995 году. За прошедшие 14 лет определена нуклеотидная последовательность десятков
геномов прокариот и эукариот и стало очевидным, что традиционные представления о филогении, базирующиеся на дарвинистических представлениях о дивергенции недостаточны для понимания генеалогии видов. Филогенетические деревья, построенные на основе молекулярного анализа гомологии отдельных генов, в частности, кодирующих рибосомальные РНК, позволяют судить о степени генетического родства организмов, но не дают однозначного ответа о происхождении и эволюционной судьбе многих генов. Дело в том, что "ветвление" таких деревьев по схеме бифуркации отражает только принцип вертикальной эволюции, в ходе которой наследуется базовый набор предковых ортологичных генов (сходных у разных организмов). В результате дупликаций, мутаций и рекомбинаций из них образуются паралогичные гены, (сходные в составе одного
генома), которые увеличивают белковый арсенал и диапазон фенотипических вариаций, отражающих усложнение клеточных систем и регуляторных механизмов. В этом суть вертикальной эволюции "вверх". Другое ключевое направление эволюционного процесса связано с горизонтальным (латеральным) переносом генов между организмами, как близкородственными, так и филогенетически отдаленными, принадлежащими даже к разным царствам.
Цель данной работы:
- рассмотреть возможный механизм эволюции прокариотического эукариотического геномов;
- рассмотреть роль и мобильных генетических элементов и горизонтального переноса генетического материала в эволюции генома;
1. Эволюция прокариотического и эукариотического геномов
1.1 Геном предполагаемого общего предка про- и эукариот
Общие принципы организации наследственного материала, представленного нуклеиновыми кислотами, а также принципы записи генетической информации у про- и эукариот свидетельствуют в пользу единства их происхождения от общего предка, у которого уже была решена проблема самовоспроизведения и записи информации на основе репликации ДНК и универсальности генетического кода. Однако геном такого предка сохранял большие эволюционные возможности, связанные с развитием надмолекулярной организации наследственного материала, разных путей реализации наследственной информации и регуляции этих процессов.
Многочисленные указания на различия в организации генома, деталях процессов экспрессии генов и механизмов ее регуляции у про- и эукариот свидетельствуют в пользу эволюции названных типов клеток по разным направлениям после их дивергенции от общего предка.
Существует предположение, что в процессе возникновения жизни на Земле первым шагом явилось образование самовоспроизводящихся молекул нуклеиновых кислот, не несущих первоначально функции кодирования аминокислот в белках. Благодаря способности к самовоспроизведению эти молекулы сохранялись во времени. Таким образом, первоначальный отбор шел на способность к самосохранению через самовоспроизведение. В соответствии с рассмотренным предположением позднее некоторые участки ДНК приобрели функцию кодирования, т.е. стали структурными генами, совокупность которых на определенном этапе эволюции составила первичный генотип. Экспрессия возникших кодирующих последовательностей ДНК привела к формированию первичного фенотипа, который оценивался естественным отбором на способность выживать в конкретной среде.
Важным моментом в рассматриваемой гипотезе является предположение о том, что существенным компонентом первых клеточных геномов была избыточная ДНК, способная реплицироваться, но не несущая функциональной нагрузки в отношении формирования фенотипа. Предполагают, что разные направления эволюции геномов про- и эукариот связаны с различной судьбой этой избыточной ДНК предкового генома, который должен был характеризоваться достаточно большим объемом. Вероятно, на ранних стадиях эволюции простейших клеточных форм у них еще не были в совершенстве отработаны главные механизмы потока информации (репликация, транскрипция, трансляция). Избыточность ДНК в этих условиях создавала возможность расширения объема кодирующих нуклеотидных последовательностей за счет некодирующих, обеспечивая возникновение многих вариантов решения проблемы формирования жизнеспособного фенотипа. [1]
1.2 Эволюция прокариотического генома
По мере совершенствования и повышения надежности главных механизмов потока информации значение избыточной ДНК в повышении выживаемости организмов снижалась. В такой ситуаций одним из возможных направлений изменения генома было уменьшение его размеров за счет утраты некодирующих нуклеотидных последовательностей. Именно так можно представить эволюционный путь, пройденный геномом современных прокариот. Одновременно в качестве механизмов, поддерживающих выживаемость этих форм, в историческом развитии закреплялось свойственное им короткое время генерации, т.е. интенсивное размножение и быстрая смена поколений (кишечная палочка делится каждые 20 мин). Перечисленные особенности хорошо сочетаются с гаплоидностью прокариот, что приводит к воспроизведению в фенотипе любой мутации.
Экспрессия 95% ДНК, относительно малые размеры генома, гаплоидность, проявление в фенотипе практически каждой мутации в сочетании с коротким временем генерации обусловливают высокую приспособленность. Вместе с тем для прокариотического типа организации не свойственны обширные и разнообразные изменения структуры. Вследствие этого описанное направление эволюции, обеспечивая высокую способность к выживанию (прокариоты существуют на Земле около 3,5 млрд. лет), является тупиковым в плане прогрессивной эволюции живых существ. [1]
1.3 Эволюция эукариотического генома
В отличие от изменений прокариотического генома преобразования генома в эволюции эукариот связаны с нарастающим увеличением количества ДНК. Это увеличение наблюдается в процессе прогрессивной эволюции эукариот . На фоне такого увеличения большая часть ДНК является ВлмолчащейВ», т.е. не кодирует аминокислот в белках или последовательностей нуклеотидов в рРНК и тРНК. Даже в пределах одного гена молчащие (интроны) и кодирующие (экзоны) участки могут перемежаться. В составе ДНК обнаруживаются высоко и умеренно повторяющиеся последовательности. Вся масса ДНК распределена между определенным числом специализированных структур тАФ хромосом. Хромосомы в отличие от нуклеоида прокариот имеют сложную химическую организацию. Эукариоты в большинстве случаев диплоидны. Время генерации у них значительно больше, чем у прокариот. Отмечаемые особенности, оформившиеся в ходе эволюции генома эукариот, допускают широкие структурные изменения и обеспечивают не только адаптивную (приспособительную), но и прогрессивную эволюцию.
Среди перечисленных выше моментов увеличение размеров генома в эволюции эукариот привлекает особое внимание. Этот процесс может осуществляться различными способами. Наиболее резко размер генома изменяется в результате полиплоидизации, которая достаточно широко распространена в природе. Она заключается в увеличении количества ДНК и хромосом, кратном гаплоидному. Достигаемое в результате состояние полиплоидии приводит к увеличению дозы всех генов и создает избыток ВлсырогоВ» генетического материала, который впоследствии видоизменяется в результате мутаций и отбора.
По-видимому, в ходе эволюции в результате накопления мутаций и дивергенции нуклеотидных последовательностей полиплоидизация сопровождалась переходом к диплоидному состоянию. Само по себе увеличение дозы генов еще не означает достижения однозначно положительного биологического результата. Об этом свидетельствует развитие в эволюции эукариот механизмов компенсации возрастающей дозы генов в ходе их экспрессии путем сокращения времени жизни в клетках зрелой РНК. Так, у тетраплоидных карповых рыб в ответ на увеличение дозы генов рРНК в молекулах рРНК соматических клеток возникают скрытые внутренние разрывы, которые приводят к преждевременному их старению и сокращению содержания в цитоплазме.
Если бы увеличение объема генома происходило только в результате полиплоидизации, то в природе должно было бы наблюдаться скачкообразное изменение его размеров. На самом деле этот процесс демонстрирует плавное увеличение содержания ДНК в геноме. Это позволяет допустить возможность других механизмов, изменяющих его объем.
Действительно, некоторое значение в определении размера генома имеют хромосомные перестройки, сопровождающиеся изменением содержания ДНК в них, такие, как дупликации, делеции и транслокации. Они обусловливают повторение, утрату некоторых последовательностей в составе хромосомы или перенос их в другие хромосомы.
Важным механизмом увеличения объема генома является амплификация нуклеотидных последовательностей, которая заключается в образовании их копий, что приводит к возникновению повторяющихся участков ДНК. Особенностью генома эукариот является наличие таких повторов в большом количестве, свидетельствующее о существенном вкладе механизма амплификации в увеличение размеров наследственного материала. Амплифицированные последовательности образуют семейства, в которых они собраны вместе (тандемная организация) или же распределяются по разным хромосомам. Конкретные изменения, приводящие к амплификации, бывают различными. Появление тандемов повторяющихся последовательностей объясняется, например, неравным кроссинговером, вследствие которого возникают многократные дупликации отдельных участков ДНК. Возможна амплификация путем вырезания фрагмента с последующей его репликацией вне хромосомы и встраиванием копий в другие хромосомы. Предполагают также амплификацию, осуществляемую путем Влобратной транскрипцииВ» ДНК на РНК с участием фермента обратной транскриптазы с последующим встраиванием копий ДНК в различные локусы хромосом.
Во всех случаях амплификация некоторой последовательности приводит к возникновению в геноме более или менее многочисленных повторов и способствует некратному увеличению его объема. Наличие таких повторов в сочетании с мутационным процессом является предпосылкой дивергентной эволюции однотипных последовательностей в пределах семейства с соответствующим изменением свойств кодируемых белков или РНК.
Ярким примером эволюционной судьбы амплифицированных нуклеотидных последовательностей являются семейства глобиновых генов, широко распространенных в природе у видов разных уровней организации. У высших позвоночных известен ряд глобиновых генов, контролирующих синтез полипептидов гемоглобина. У человека в геноме имеется восемь активных глобиновых генов, образующих два семейства.
Семейство генов, определяющих синтез а-глобинов, содержит два тАУглобиновых гена, активно функционирующих в эмбриогенезе, и два
тАУглобиновых гена, активно функционирующих в эмбриогенезе, и два 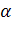 -глобиновых гена, которые экспрессируются у плода и взрослого человека. Это семейство генов располагается в 16-й хромосоме в следующем порядке: 5' тАФ
-глобиновых гена, которые экспрессируются у плода и взрослого человека. Это семейство генов располагается в 16-й хромосоме в следующем порядке: 5' тАФ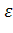 2 тАФ1тАФ 2тАФ
2 тАФ1тАФ 2тАФ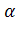 1тАФ3'. Семейство генов, определяющих синтез
1тАФ3'. Семейство генов, определяющих синтез 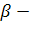 глобинов, расположенное в 11-й хромосоме, содержит
глобинов, расположенное в 11-й хромосоме, содержит 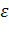 тАУглобиновый ген эмбриона, два сходных
тАУглобиновый ген эмбриона, два сходных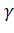 тАУглобиновых гена плода
тАУглобиновых гена плода  и
и 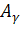 , малый
, малый  - и большой
- и большой 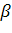 -глобиновые гены взрослых: 5' тАФ тАФ тАФ тАФ
-глобиновые гены взрослых: 5' тАФ тАФ тАФ тАФ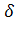 тАФ
тАФ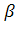 тАФ 3'.
тАФ 3'.
Изучение гомологии продуктов указанных генов и генов миоглобина у разных видов организмов позволило предположить общность происхождения этих семейств. Вероятно, около 1100 млн. лет назад произошла дупликация гена-предшественника, давшая начало гемоглобиновым и миоглобиновым генам. Позднее, около 500 млн. лет назад, на ранней стадии эволюции позвоночных произошла дупликация, давшая начало двум ( и ) семействам глобиновых генов, сопровождавшаяся транслокацией. Примерно 200 млн. лет назад очередная дупликация привела к возникновению в семействе тАУглобиновых генов генов тАУглобинов плодов и взрослых. Около 100 млн. лет назад произошло образование тАУ 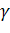 -глобиновых генов и, наконец, 40 млн. лет назад появились тАУ и тАУглобиновые гены.
-глобиновых генов и, наконец, 40 млн. лет назад появились тАУ и тАУглобиновые гены.
Семейства тАУ и тАУглобиновых генов организованы в генные кластеры, возникшие, вероятно, в результате тандемной дупликации генов. В составе указанных кластеров наряду с активно функционирующими на разных стадиях онтогенеза генами обнаружены неактивные, или псевдогены. Последние возникли, вероятно, в результате появления в них изменений, несовместимых с возможностью их экспрессии. В семействе тАУглобиновых генов содержится два псевдогена:  и
и 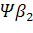 . В -семействе имеется один псевдоген. Дивергенция амплифицированных последовательностей с образованием разных генов или их семейств обусловлена накоплением в них различных изменений в виде замен оснований или других генных мутаций. О гомологии глобиновых генов обоих семейств свидетельствует наличие во всех активных глобиновых генах позвоночных двух интронных участков, занимающих в них строго одинаковое положение. Такую же организацию имеют и псевдогены
. В -семействе имеется один псевдоген. Дивергенция амплифицированных последовательностей с образованием разных генов или их семейств обусловлена накоплением в них различных изменений в виде замен оснований или других генных мутаций. О гомологии глобиновых генов обоих семейств свидетельствует наличие во всех активных глобиновых генах позвоночных двух интронных участков, занимающих в них строго одинаковое положение. Такую же организацию имеют и псевдогены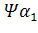 у человека,
у человека, 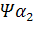 у кролика. Однако в
у кролика. Однако в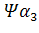 псевдогене мыши в ходе эволюции оба интрона оказались точно вырезанными.
псевдогене мыши в ходе эволюции оба интрона оказались точно вырезанными.
Результатом амплификации небольших последовательностей ДНК в пределах функциональной единицы является удлинение гена, при котором из простых генов могут возникать более сложные. Это может происходить за счет тандемных дупликаций. Например, в генах, кодирующих вариабельные участки иммуноглобулинов мыши, последовательности из 600 п.н. образуются в результате 12 тандемных повторов исходной предковой последовательности в 48 п.н. Другим примером удлинения гена посредством тандемных дупликаций служит ген коллагена , который у курицы состоит из 34 000 т.н. и содержит больше 50 экзонов. Длина таких участков во всех случаях кратна девяти нуклеотидным парам. Эволюция этих экзонов, очевидно, шла от гипотетического исходного строительного блока длиной в 54 пары нуклеотидов.
, который у курицы состоит из 34 000 т.н. и содержит больше 50 экзонов. Длина таких участков во всех случаях кратна девяти нуклеотидным парам. Эволюция этих экзонов, очевидно, шла от гипотетического исходного строительного блока длиной в 54 пары нуклеотидов.
Таким образом, амплификация нуклеотидных последовательностей, происходившая в процессе эволюции генома, обеспечивала не только его количественное увеличение, появление семейств генов, но и создавала предпосылки для накопления в них изменений, дивергенции генов, увеличения разнообразия контролируемых ими продуктов. [1]
2. Роль и мобильных генетических элементов горизонтального переноса генов в эволюции генома
2.1 Мобильные генетические элементы, их свойства
Мобильные генетические элементы (МГЭ, подвижные элементы, транспозируемые элементы, транспозоны и т.д.) повсеместно распространены в живой природе от плазмид, фагов и бактерий до высших животных и растений. Будучи нестабильными по своей локализации в геномах, они создают мощный источник изменчивости генов, систем их управления и геномов. Будучи сами последовательностями нуклеотидов, они тоже подвержены эволюции. Поэтому МГЭ выступают и как факторы эволюции содержащих их геномов, и как эволюционирущие объекты.
В настоящее время МГЭ найдены у бактерий (включая их фагов и плазмид), низших грибов, насекомых, растений, животных и многих других объектов. Число известных семейств МГЭ, вероятно превышает 100. У хорошо изученных объектов найдены многие десятки семейств МГЭ: например у дрозофилы их число, вероятно, достигает 50. Некоторые семейства относятся к умеренным повторам, имея десятки-сотни копий на геном, другие - к повторам высокой множественности (Alu у человека - 5тАФ6 млн копий на геном). В сумме МГЭ различных семейств могут составлять значительную часть генома (до 10% у дрозофилы). МГЭ нестабильны, т.е. с определенной вероятностью способны к транспозициям и исключению из отдельных позиций генома (табл.1).
Таблица 1. Некоторые свойства МГЭ (Хесин, 1984; Fennegan,1985)
| МГЭ | Частота транспозиций на позицию, за поколение | Количество копий МГЭ в геноме | МГЭ | Частота транспозиций на позицию, за поколение | Количество копий МГЭ в геноме |
| FB | 10-3 | 30 | Te | 10-3 | 10 |
| P | 10-2-10-5 | 30-50 | MU | 10-1-10-3 | 10-60 |
| Ty | 10-7 | 30-35 | MDG | 10-2-10-5 | 10-50 |
МГЭ способны к воспроизведению в клетке либо через репликацию ДНК, либо через прямую и обратную транскрипцию (ретротранспозоны).В случаях, когда МГЭ не содержит генов, выполняющих клеточных функции, их часто считают "эгоистическими ДНК". Транспозиции обычно связаны с размножением копий МГЭ. В своей структуре МГЭ содержат гены транспозиции (ферментов-транспозаз - Тп-3, Тп-5 ) и др., ревертаз - ретропозоны (Ананьев, 1989). Поэтому фактически они являются отдельными репликонами. В некоторых случаях синтез транспозазы репрессируется при избыточной ее концентрации по механизму отрицательной обратной связи (Тп-3), Р-фактор дрозофилы). МГЭ содержат также разнообразные функциональные сайты - знаки пунктуации и управления (промоторы, терминаторы, операторы, репликаторы, энхансеры, регуляторные сайты теплового шока), которые существенны для окружающих участков генома. Инсерции МГЭ в кодирующие зоны генов приводят к нарушению или резкому изменению их функций. Это связано с прямым нарушением генов и с влиянием знаков пунктуации (промоторов, терминаторов и др.,) на процессы считывания. Доля таких мутаций особенно велика у прокариот, которые имеют высокую плотность кодирования информации в геноме. Инсерции МГЭ в некодирующие области (спейсеры, интроны, фланговые участки др.) приводят к более "мягким" последствиям: усилению или ослаблению активности близлежащих генов, изменению их регуляции и т.п. Такие последствия преобладают у высших эукариот, у которых кодирующая часть генома составляет ~3-5%. Показано также, что среди видимых мутаций у дрозофилы и других объектов наиболее значительную долю составляют не замены нуклеотидов, а именно инсерции МГЭ (табл. 2).
Таблица 2 Доля спонтанных мутаций, вызванных инсерциями МГЭ, в различных локусах Drosophilla melanogaster (Sankaranarayanan, 1988; McDonald, 1989)
| Локус | Доля инсерций МГЭ среди видимых спонтанных мутаций | МГЭ (число мутаций) |
| V (vemillion) | 4/5 | mg-2-(3), BI04 (1) |
| ct (cut) | 28/28 | mdg-4 (27), copia (1) |
| ry (rosy) | 3/5 | calypso (1), BI04 (1) |
| f (forked) | ¾ | mdg-4 (3) |
| su (s) | 5/7 | mdg-4 (5) |
| Bx (Beadex) | 4/4 | mdg-4 (2) ,BI04 (2) |
| bx (bithorax) | 8/9 | mdg-4 (7) ,BI04 (2) |
| sc (scute) | 2/2 | mdg-4 (2) |
| Antr (Antennapedia) | 2/5 | BI04 (2) |
2.2 Отбор, температурные воздействия и динамика рисунка локализации МГЭ в геноме
Гибридизация in situ ДНК МГЭ с политенными хромосомами дрозофилы была тем методом, который позволил исследовать популяционную динамику рисунков локализации МГЭ. Гвоздев, Кайданов и др. впервые обнаружили, что отбор по количественному признаку половой активности самцов дрозофилы приводил к направленному изменению рисунка локализации copia-подобных МГЭ. Сходные явления были обнаружены также в других системах МГЭ дрозофилы.
Васильева и др. (1987) селекционировали лабораторную популяцию D. melanogaster с прерванной радиальной жилкой крыла (олигогенная мутация radius incompletus тАФ ri). Контрольная популяция riC имела проксимальный и дистальный фрагменты (табл. 3).
Таблица 3. Средняя длина проксимального (Х1) и дистального (Х2) фрагментов радиальной жилки крыла у самок различных линий дрозофил (генеалогию линии см. на рис. 1)
| линия | год начала культивирования линии | поколение | Х1 | Х2 |
| riC | 1974 | 220 | 1.91÷0.05 | 0.39÷0.07 |
| riSN | 1974 | 220 | 0.50÷0.02 | отсутствует |
| riTII3 | 1979 | 140 | 1.05÷0.05 | отсутствует |
| riTII49 | 1979 | 140 | 2.45÷0.09 | 1.47÷0.08 |
| riSP | 1982 | 40 | 3.80÷0.02 | 2.20÷0.03 |
В результате 70 поколений минус-отбора жилка была почти полностью редуцирована, и далее фенотип устойчиво воспроизводился без дальнейшего отбора в течение более 300 поколений (линия riSN). В результате 40 поколений плюс-отбора жилка была полностью восстановлена, фенотип далее тоже воспроизводится без дальнейшего отбора многие десятки поколений (линия riSP). В табл. 3 приведены количественные итоги селекции. Сравнительный анализ рисунков локализации МГЭ mdg-1, mdg-2 и copia показал (Васильева и др., 1987; Забанов и др., 1990), что все они имеют в контрольной линии совершенно различные рисунки локализации. В результате отбора по признаку в любом из двух направлений происходили характерные изменения рисунков, хорошо воспроизводимые в повторных экспериментах. Результаты изменения признаков и рисунков локализации МГЭ достаточно устойчиво воспроизводились в отселекционированных линиях. На рис. 1 изображено генеалогическое дерево этих и других исследованных линий.
Куколки дрозофил контрольной популяции (riC) были подвергнуты температурному воздействию: ступенчатому изменению температуры культивирования тАФ 29В° -В» 18В° С. Фенотипы обработанных особей оказались чувствительными к этому фактически стрессовому воздействию. В наибольшей степени это проявилось при обработке в течение двух коротких периодов куколочной стадии. Опуская детали, укажем, что главным эффектом было изменение экспрессии признака в последующих поколениях, которое устойчиво наследуется свыше 200 поколений. Температурная обработка в 1-й чувствительный период (113 В±5 ч.) приводила к существенной редукции количественного признака в последующих поколениях, обработка во 2-ой чувствительный период (149 В± 5 ч.) тАФ к почти полному восстановлению жилки. Соответствующие "температурные" линии были названы riТIIЗ и riТI49 (см. табл. 3 и рис. 1).
Генетический анализ "селекционных" и "температурных" линий показал, что полигенные факторы, влияющие на количественный признак, распределены по всем хромосомам дрозофилы (Васильева и др., 1987; Забанов и др., 1990).
Гибридизация in situ тех же copia-подобных элементов с полигенными хромосомами личинок этих линий выявила вполне определенные, существенные и воспроизводимые изменения рисунков локализации по сравнению с контрольной линией (riC). Таким образом, разовое температурное стрессовое воздействие вызывает множественные транспозиции copia-подобных МГЭ и изменения полигенов, сопровождаемые также изменениями фенотипа. Укажем, что по фенотипу полученные "температурные" линии попарно сходны с "селекционными": riSN с пТПЗ, riSP с riTI49 (см. табл. 3). В ряде изогенных линий дрозофилы множественные перемещения МГЭ обнаружены непосредственно в следующем поколении (Колесникова и др., 1991).
Рисунки локализации МГЭ по сегментам цитологической карты полигенных хромосом представляют собой линейные последовательности бинарных символов. Их можно сравнивать между собой теми же способами, что и последовательности мономеров. В каждой из пяти линий были взяты по 7тАФ9 личинок. Для каждой пары последовательностей строились расстояния по Хеммингу, матрица таких расстояний была использована для построения дерева сходства. На рис. 2 изображено такое бескорневое дерево, построенное методом UPGMA для рисунков mdg-2.
Основной результат такого анализа состоит в том, что линии с наиболее сходными рисунками локализации МГЭ оказались также наиболее близкими и по количественным проявлениям признаков: riSN и пТПЗ, riSP и riTI49. Еще раз подчеркнем, что сходство по признаку и по рисунку локализации МГЭ проявили линии, полученные принципиально разными путями: "селекционные" тАФ путем длительного жесткого отбора по фенотипу, "температурные" тАФ путем разовой температурной обработки в чувствительные периоды развития. Этот факт свидетельствует о том, что рисунок локализации МГЭ и экспрессия признака связаны между собой причинно (генотип тАФ фенотип).
Убедимся, что с точки зрения популяционных механизмов тАФ генетического дрейфа, накопления транспозиций и др.тАФ эта связь не случайна. Во-первых, она воспроизводима при повторном выводе "температурных" линий в 1979,1982,1985,1986 гг. Во-вторых, роль генетического дрейфа в возникновении различий между линиями можно считать малой. При культивировании всех линий выполнялся комплекс антидрейфовых мероприятий, препятствующих случайной фиксации или потере вариантов локализации МГЭ (..) т.е. случайные флуктуации частот нивелировались, а фиксация становилась маловероятной. В-третьих, мы имели для сравнения два независимо построенных дерева тАФ генеалогическое дерево линий (см. рис. 1), отражающее реальный процесс их выведения, и дерево сходства линий (рис. 2), отражающее степень их близости по рисункам локализации МГЭ. Сравнения этих деревьев показывает, что они совершенно различны. Это значит, что в процессе выведения и микроэволюции этих линий были нарушены условия, необходимые для того, чтобы дерево сходства дивергировавших последовательностей отражало реальную топологию дерева эволюции. В-четвертых, имеется ряд факторов, указывающих на "взрывообразный" характер транспозиций МГЭ в "температурных" линиях. По данным Колосниковой и др. (1991), в изогенных линиях, выведенных из контрольной линии пС, в следующем поколении после теплового шока найдены множественные транспозиции mdg-2. Юнакович и др. , использовав блотинг по Саузерну и зонды на присутствие copia-подобных МГЭ, показали, что после теплового шока в следующем поколении происходили массовые перемещения МГЭ. С другой стороны, и в "селекционных" линиях отбор происходил весьма неравномерно и занял лишь часть времени микроэволюции, а затем был прекращен, т.е. и здесь нельзя говорить о постоянстве скорости фиксации транспозиций (Васильева и др., 1987). Именно эти особенности являются причиной несовпадения двух построенных деревьев (рис. 1, 2). Заметим, что этот случай является одним из наиболее наглядных примеров того, что дерево сходства последовательностей не всегда изоморфно отображает реальный процесс эволюции.
Наконец, в-пятых, отметим, что сходство линий по рисункам локализации МГЭ может быть либо "остаточным", поскольку все линии имеют общее происхождение (см. рис. 1), либо вновь приобретенным, если спектры изменений у линий сходны, неслучайны. Сравнение спектров отличий четырех дочерних линий от контрольной (riC) показывает, что наиболее сходные по фенотипу линии имеют наиболее сходные спектры изменений локализации mdg-2. Так, линии riSN и riT 113 имеют 27 общих изменений из 33тАФ35 по сравнению с riC; линии riSP и riT 149 имеют 17 общих изменений из 23тАФ30 замен. Следовательно, подавляющая часть спектров изменений в высокой степени не случайна по своему положению канализована.
Многие другие copia-подобньге МГЭ обладают подобными же свойствами. Рисунки локализации mdg-1, mdg-3, copia почти полностью различны, но деревья сходства рисунков похожи друг на друга. В то же время, рисунки МГЭ вовлекаются в отбор по многим различным количественным признакам. Поэтому можно полагать, что мы имеем дело с геномной системой влияния МГЭ на проявление различных генов, чувствительной к стрессовым внешним воздействиям и отвечающей на селекцию. [4]
2.3 Гипотеза: эволюционная роль системы МГЭ в геномах эукариот
Суммируя вышеизложенное, можно констатировать, что система МГЭ эукариотического генома обладает, по крайней мере, следующими общими функциями: 1) является источником инсерционной изменчивости генов; 2) влияет на проявление количественных и качественных признаков; 3) откликается изменением рисунков локализации многих МГЭ на отбор по признакам; 4) откликается на внешние стрессорные воздействия, в частности тАФ температурные, вспышками транспозиционной изменчивости. Такими свойствами в разной степени обладают МГЭ различных объектов тАФ дрожжей, дрозофилы, растений, млекопитающих.
Механизм инсерционной изменчивости достаточно ясен. Отметим только, что активация транспозиций разными путями (дисгенное скрещивание а РтАФМ системе (МсКау, 1988), температурное воздействие в системе copia-подобных МГЭ должна усиливать эти компоненты изменчивости. Укажем также на существование "транспозиционных взрывов" в отдельных генеративных клетках дрозофилы, возникающих после генетического воздействия или спонтанно.
Механизм влияния МГЭ на экспрессию генов эукариот требует пояснений. В целом МГЭ содержат широкое разнообразие регуляторных сайтов, среди которых наиболее интересны энхансеры и регуляторные сайты теплового шока. Известно, что энхансеры способны в десятки и сотни раз усиливать транскрипцию соседних генов на расстоянии до 5000 н.п. методом контекстного анализа показали, что в структуре многих МГЭ (в том числе тАФ mdg-1, mdg-2, mdg-4, copia) имеются энхансеро-подобные сайты.
Регуляторные сайты теплового шока (РСТШ) присутствуют в 5'-областях перед началом транскрипции эукариотичкеских генов, проявляющих эффект теплового шока. Размер их ~14 н.п., расстояние до блока Хогнесса sS 200 н.п. Восстановлен консенсус РСТШ, характерный для таких далеких форм, как дрожжи, дрозофила, лягушка, человек: CNNGAANNTTCNNG (NтАФлюбой нуклеотид). Здесь 8 консервативных позиций. В некоторых случаях РСТШ имеют другую структуру. Чем ближе РСТШ к блоку Хогнесса, тем эффективнее активация транскрипции тепловым шоком (Bienc, 1985). РСТШ можно рассматривать как энхансеры с позитивной регуляцией.
Показано, что активация транскрипции МГЭ copia у дрозофилы и DIRS1 у дрожжей при тепловом шоке связана с наличием РСТШ в их терминальных повторах). Капитонов и др. (1987) методом контекстного анализа обнаружили РСТШ-подобные сайты в семи из 13 исследованных ими МГЭ: mdg-1, mdg-4, hobo, P дрозофилы, BS1 и CIN1 кукурузы и в ретровирусе EV1 курицы. Все они находились на расстоянии ^ 150 н.п. от начала транскрипции и имели неслучайную гомологию либо с известными РСТШ, либо с их консенсусом. В mdg-2 и в МГЭ H.M.S. Beagle дрозофилы этот вариант РСТШ не обнаружен, но это не исключает возможного присутствия других РСТШ. Сходные результаты независимо получены также Макдональдом и сотрудниками. Интересно, что РСТШ найден также в последовательности секвенированного ретровируса HTLVтАФIII (вирус СПИД). В некоторых МГЭ найдены РТСШ-подобные сайты на расстоянии > 200 н.п. от блока Хогнеса. Отметим, однако, что наличие РТСШ-подобного сайта еще не гарантирует его функционирование.
Прямое доказательство индукции транспозиций copia-подобного МГЭ mdg-2 при помощи теплового шока получено Колесниковой и др. (1991). В изогенной линии, самцы которой были подвергнуты тепловому шоку, транспозиции были найдены в следующем поколении, причем вероятности транспозиций возросли более чем на два порядка величин.
Таким образом, появление дополнительного энхансера вблизи функционального гена в результате инсерции несущего его МГЭ способно резко активизировать этот ген. Инсерция МГЭ, содержащего РСТШ, способна подчинить соседние гены системе теплового шока и сделать их чувствительными к стрессорным факторам окружения. В связи с этим можно сформулировать гипотезу о роли МГЭ в ответе на отбор по признакам и в температурных эффектах.
На основе имеющихся фактов (см. выше) необходимо постулировать, что рисунки локализации МГЭ являются существенной компонентой генетического механизма детерминации количественных признаков (Vasilyeva et al., 1985; Ratner, Vasilyeva, 1989). В общем случае эти системы содержат: 1) олигогены (гены главного эффекта), необходимые для формирования признака; 2) полигены, каждый из которых не необходим для формирования признака, но в совокупности они могут существенно изменить его экспрессию; 3) МГЭ, которые модифицируют, усиливают действие олигогенов и полигенов, вблизи которых они локализованы. Рисунок локализации МГЭ в каждом случае относительно стабилен; позиции, доступные для инсерции, вероятно, мечены на длительный срок "молекулярной памятью", существование которой в частном случае показано Мизрохи и др. (1985); возможно, что список доступных позиций зависит от компактизации хромосом и других явлений ядерно-хромосомного уровня. Рисунок локализации МГЭ распределяется по этим специфическим позициям (см. параметры n и m в п. 2). Скорее всего, рисунок МГЭ более или менее случайно наложен на топографию локализации олигогенов и полигенов и активирует их.
В рамках концепции лимитирующих факторов необходимо также предположить, что влияние как полигенов, так и МГЭ проявляется фенотипически только в тех случаях если они действуют на лимитирующие олигогены. Тогда их эффект оценивается отбором: вариабельность рисунков будет создавать вариабельность лимитирующих признаков, отбор по признакам будет приводить к отбору полигенов и рисунков модифицирующих их МГЭ. Определенный вклад в изменчивость признаков будут вносить также новые редкие транспозиции МГЭ. Ясно, однако, что селективное изменение рисунков не будет равномерным, поскольку это не нейтральный процесс фиксации перемещений МГЭ.
Для объяснения температурных эффектов предполагается, что стрессовое температурное воздействие (тепловой шок, ступенчатое изменение температуры и др.) индуцирует вспышку множественных у отдельных особей и массовых в популяции транспозиций МГЭ, которые выявляются уже в следующем поколении (Колесникова и др., 1991) и приводят к наследуемому изменению количественных и качественных признаков. Неслучайный характер температурно-индуцированных транспозиций можно связать с существованием каких-то достаточно жестких ограничений на локализацию этих транспозиций в геноме. Имеется в виду, что в рамках группы выделенных сайтов, в принципе доступных для инсерций МГЭ, т.е. обладающих гомологией к ним, имеются дополнительные ограничения на транспозиции, связанные со свойствами хромосом или клеточного ядра и изменяющиеся в ходе онтогенеза. Эти ограничения должны быть различны на разных этапах онтогенеза (в чувствительные периоды). Сходство фенотипов "селекционных" и "температурных" линий сопровождается сходством спектров перемещения МГЭ. Это возможно только в том случае, если спонтанные неиндуцированные транспозиции в "селекционных" линиях подчиняются таким же ограничениям, что и индуцированные в "температурных" линиях.
Последствия температурных воздействий могут быть
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)