Липиды центральной нервной системы и структура клеточных мембран
Липиды являются не только структурными компонентами ЦНС, но и важнейшими участниками функциональной активности. Головной мозг характеризуется высоким содержанием липидов. Мозг содержит уникальные мембранные структуры тАУ миелиновые оболочки, которые имеют самое высокое содержание липидов по сравнению с другими тканями или субклеточными структурами, за исключением адипозной ткани. Для ЦНС характерно и наибольшее структурное разнообразие липидов по сравнению с мембранами других органов.
Липидный состав нервной ткани практически постоянен и остается неизменным даже под влиянием внешних факторов, которые меняют липвдный состав висцеральных органов и плазмы. Это тАУ следствие защищенности ЦНС от различных внешних воздействий. Изменение липидного состава нервной ткани рассматривается обычно как патология, хотя при этом следует помнить, что существенные изменения в липидном составе нервной системы происходят в период развития.
Вся сложнейшая деятельность нервной ткани опосредуется через мембраны, в формировании и функционировании которых липиды принимают непосредственное участие.
В клетках нервной системы представлено несколько типов высокоспециализированных мембран: соматические мембраны мульти- и униполярных нейронов, мембраны дендритов, миелинизированных и немиелинизированных аксонов, аксонного холмика, где генерируется потенциал действия, мембраны рыхлого и компактного миелина, мембраны синаптических пузырьков, пре- и постсинаптические мембраны, мембраны макро- и микроглии. Возбудимость этих мембран колеблется в широких пределах от высоковозбудимых до относительно устойчивых мультимембранных структур миелина. В составе, строении и функционировании мембран нервной ткани еще очень много неясного. Для того чтобы раскрыть надмолекулярную организацию этих мембран, надо иметь достаточно полное представление об их липидном и белковом составе. Однако исследователи пока не владеют этими сведениями в полной мере, хотя ряд важных закономерностей уже намечен.
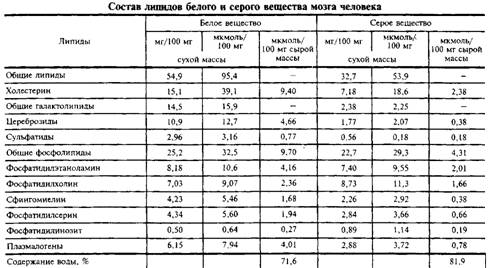
Липидный состав серого и белого вещества мозга человека представлен в табл. 1, а различных клеток мозга тАУ в табл. 2. Видно, что липидный состав белого вещества ближе к миелину, а серое вещество содержит меньше типичных миелиновых ли-пидов, но относительно больше ганглиозидов.
Сравнивая молярное содержание основных классов липидов в специализированных клетках мозга, можно видеть, что оли-годендроглия и миелин наиболее обогащены цереброзидами, а нейроны и астроглия имеют более высокое содержание фосфолипидов. Это лишний раз подтверждает, что плазматические мембраны совершенно отличны от миелина.
Состав фосфолипидов обогащенных фракций нейронов и нейроглии коры мозга крысы представлен в табл. 3.
Чем более анатомически дифференцированно подходить к нервной ткани, тем больше различий обнаруживается в липидном составе, поскольку функционально различные нейрональ-ные и глиальные клетки имеют своеобразный липидный состав.
В состав большинства липидов входят жирные кислоты. В мозге они гораздо разнообразнее, чем в других тканях. Это намного увеличивает число индивидуальных липидов мозга. Содержание жирных кислот в головном мозге гораздо выше, чем в других органах, и составляет примерно 20тАУ25% в расчете на сухую массу ткани. Разнообразие жирных кислот в этом органе поразительно. Применение газожидкостной хроматографии позволило продемонстрировать наличие в головном мозге более 50 жирных кислот с длиной цепи от 12 до 26 углеродных атомов, среди которых найдены насыщенные, ненасыщенные, нормальные, гидроксизамещенные, нечетные и др. Ненасыщенные кислоты мозга могут содержать от 1 до 6 двойных связей. Особенностью, липидов мозга является относительно большое содержание длинноцепочечных полиеновых кислот 20:4, 22: 5, 22:6.
Отдельные классы и фракции липидов мозга характеризуются своим набором жирных кислот. Имеет место также определенная специфичность жирнокислотного состава в лип идах разных отделов мозга, разных типов его клеток, субклеточных структур. Иллюстрацией этого могут служить данные табл. 4, где приведен жирнокислотный состав фосфолипидов синаптосом и миелина тАУ двух разных типов мембранных структур ЦНС,
Состав липидов основных типов нервных клеток мозга крысы
| Липиды | Нейроны | Астроглия | Олигоден-дроглия | Миелин |
| Холестерин | 6,610 | 14,100 | 10,800 | 54,900 |
| Цереброзиды | 0,513 | 0,689 | 2,610 | 22,000 |
| Сульфатиды | 0,090 | 0,142 | 0,472 | 2,890 |
| Общие фосфолипиды | 22,400 | 35,600 | 23,400 | 41,800 |
| Ганглиозиды | 0,223 | 0,582 | 0,239 | 0,0453 |
| Молярное отношение: холестерин тАУ цереброзиды-фосфолипиды | 1:0,075:3,5 | 1:0,05:2,5 | 1:0,25:2,2 | 1:0,40:0,76 |
| Фосфолипиды | Нейроны | Нейроглия |
| Лизофосфатидилхолин | 3,9 | 1,9 |
| Фосфатидилхолин | 46,1 | 46,9 |
| Сфингомиелин | 6,7 | 9,5 |
| Фосфатидилсерин | 9,1 | 7,1 |
| Фосфатидилинозит | 7,7 | 5,9 |
| Фосфатидил этанол а мин | 25,1 | 24,9 |
| Фосфатидная кислота | 1,8 | 3,6 |
Содержание индивидуальных фосфолипидов в коре мозга крысы резко различающихся по своему происхождению и функциям.
В синаптосомах велико содержание жирных кислот тАУ С 22:6, а в миелине высок процент моноеновых кислот тАУ 18:1. Возможно, что высокое содержание докозагексаеновой кислоты в синаптосомах необходимо для активного транспорта ионов, так как активность Na+, К+-АТФазы в них зависит от присутствия полиеновых кислот в фосфолипидах. В мозге имеются регуля-торные механизмы, поддерживающие степень ненасыщенности и специфичность жирнокислотного состава в липидах.
Состав жирных кислот фосфолипидов сннаптосомальных и миеляновых мембран коры мозга обезьяны
| Шифр жирной кислоты | Фосфатид ил- |
| холи н | этанол амин | серин+монофосфо-инозитид |
| синапто-сомы | миелин | синапто-сомы | миелин | синапто-сомы | миелин |
| 16:0 | 50 | 33,1 | 7,4 | 4,9 | 3,5 | 2,6 |
| 18:0 | 12,4 | 17 | 25,5 | 15,9 | 44,3 | 43 |
| 18:1 | 27,2 | 42,3 | 12,1 | 33,2 | П, 4 | 38,5 |
| 20:1 | 0,7 | 0,9 | 1,6 ■ | 9,3 | - | 2,5 |
| 20:4 | 3,8 | 3,2 | 10,1 | 11,6 | 8,3 | 6,3 |
| 22:4 | 0,8 | 0,6 | 6,4 | 13,1 | 3,5 | 3,7 |
| 22:6 | 3 | 2,3 | 24,9 | 10,6 | 26,9 | 2,9 |
Изменение жирнокислотного состава приводит к нарушению функциональной деятельности мозга.
1. РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
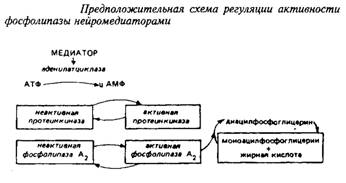
В мембранах головного мозга имеет место цикл деацилирование тАУ реацилирование, при котором происходит замена жирных кислот в молекуле фосфолипидов, в то время как другие компоненты молекулы остаются неизменными. Этот ацилобменный механизм является особенно важным для включения тех или иных жирных кислот во второе положение остатка глицерина, и его рассматривают как средство локального регулирования физических и функциональных свойств мембран. Существенную роль играет и переход диацильных форм фосфолипидов в моноацильные и обратно. Все это оказывает влияние на такие мембранные процессы, как проницаемость для различных веществ, транспорт ионов и т.д.
Ацилобменные реакции имеют прямое отношение ко многим процессам, влияя на активность ряда ферментов, на синтез простагландинов и чувствительность фоторецепторов. Некоторые исследователи связывают ферментативное деацилирование тАУ реацилирование с эффектом синаптической передачи. Так, под влиянием норадреналина в синаптосомах происходит активирование фосфолипазы А2, отщепляющей жирную кислоту во втором положении глицерофосфолипида. Таким образом, нейромедиатор модифицирует обмен фосфолигавдов в синаптических мембранах путем вовлечения в этот процесс реакций деацилирования. Предложена следующая схема регуляции активности ацилобменного цикла нейромедиаторами.
2. ОРГАНИЗАЦИЯ ЛИПИДОВ В МЕМБРАНЕ
Образование липидных молекул в ходе эволюции и выбор именно этих молекул в качестве строительных блоков мембран сыграли решающую роль в возникновении жизни. Липидам принадлежит жизненно важная роль в клетке. Следующие особые физико-химические свойства липидов определяют их роль в построении мембран:
1. Сочетание гидрофильных и липофильных свойств в структуре одной молекулы, их амфифильность.
2. Способность липидов четко ориентироваться на границе раздела фаз, так что полярные группы направлены в водные среды, а неполярные экранированы от них.
3. Способность липидов самопроизвольно упаковываться в прочные, плотные мономолекулярные слои или пленки, устойчивые к сжатию. Плотность такой упаковки зависит от рН, температуры и молекулярной организации липидов. Такие плотные слои создают определенный барьер для диффузии молекул.
4. Способность липидов агрегировать в хорошо упорядоченные сферические, цилиндрические, ламеллярные мицеллы. В мицеллах липиды ориентированы таким образом, чтобы максимальное число полярных групп находилось в контакте с водой, а гидрофобная часть была максимально удалена от контакта с ней.
Способность липидов образовывать прочные мономолекулярные слои лежит в основе молекулярной организации мембран. Более 60 лет назад было высказано предположение, что в основе мембран лежит бимолекулярный слой липидов.
В бимолекулярном липидном слое гидрофобные цепочки молекул липидов направлены друг к другу и внутренность бислоя совершенно гидрофобна, а гидрофильные части образуют поверхности внутреннего и внешнего монослоев, открытые для разнообразного рода взаимодействий.
Липидный состав мембран нервной ткани и распределение липидов по слоям генетически детерминированы. Наружный и внутренний монослои липидов характеризуются планарной и поперечной микрогетерогенностьюу что создает асимметричность мембран. Существует несколько механизмов, поддерживающих асимметричное распределение липидов в мембране. Один из них связан с термодинамической вероятностью размещения липид-ных молекул с учетом их стереоконфигурации, заряда и гидратации полярных групп. Так, основная часть фосфатидилхолина, сфингомиелина, полифосфоинозитидов, холестерина, церебро-зидов и сульфатидов локализована в наружном слое, а амино-фосфолипиды находятся во внутреннем, цитоплазматическом слое. Неодинакова степень ненасыщенности монослоев: во внутреннем обнаруживается 2/3 двойных связей в жирных кислотах липидов, а в наружном тАУ только 1/3.
Асимметрия бислоя является фактором, обеспечивающим создание градиента кривизны, складок, сморщиваний, отшнуровки части мембраны в виде везикул что существенно для обеспечения межклеточных взаимодействий.
Другой механизм поддержания асимметрии бислоя реализуется за счет различий ионного состава вне- и внутриклеточной среды, что вносит вклад в создание и поддержание изгибов мембраны.
Асимметрия бислоя обеспечивается также ферментами ли-пидного обмена, к ним прежде всего относятся липазы, ферменты обмена холестерина и метилазы фосфатидилэтанолами-на. Метилирование фосфатидилэтаноламина с превращением его в фосфатидилхолин осуществляется в два этапа и происходит в разных слоях липидкого матрикса. Образование мономе-тилфосфатидилэтаноламина под влиянием метилтрансферазы I осуществляется во внутреннем слое, где и локализован фермент. Монометил фосфатидилэтаноламин переходит из цитоплаз-матического слоя на внешний, где под действием метилтрансферазы II завершается его превращение в фосфатидилхолин. Фактически осуществляется так называемый ферментативный флип-флоп.
Этот транслокационный процесс меняет жидкостность мембраны и рассматривается как фактор, стимулирующий функционально важные процессы в мембране: связывание рецепторов с лигандами, Са* тАУ вызванное освобождение медиаторов из си-наптических окончаний, активирование ЛТФаз.
Асимметричность билипидного слоя может поддерживаться транспортом липидов: спонтанным, везикулярным или с участием липидпереносящих белков. Липидпереносящие белки различной степени специфичности Влстоят на стражеВ» асимметрии мембран, перенося липиды только в наружный или только во внутренний слой. Перенос липидных молекул осуществляется в виде комплексов с белками-переносчиками.
3. ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
Строгая организованность липидного слоя мембраны не лишает его большой динамичности, которая возникает из-за передвижения липидных молекул в пределах мембраны, т.е. за счет интрамолекулярных движений липидов в пределах бислоя. Известно по крайней мере четыре типа интрамолекулярных движений липидов в пределах мембраны: латеральная диффузия, вращательная диффузия, вертикальные колебания и упоминавшийся выше так называемый флип-флоп.
Для большинства липидов скорость латеральной диффузии ощутима. Коэффициент латеральной диффузии для липидов Ю тАУ см/с, а для белков намного ниже тАУ 10~ ш/с. Вращательная диффузия молекул осуществляется также легко. Скорость же флип-флопа очень низка. Особенно медленно флип-флоп происходит в чисто липидных везикулах. Даже в присутствии липидпереносящих белков перемещение из наружного слоя во внутренний занимает более 4 часов, а перемещение в обратную сторону тАУ более 10 часов. Не ускоряет флип-флоп повышение температуры до 80В°С Это движение фосфолипидов усиливается под влиянием окисленных липидов, лизолецити-на. Как правило, холестерин подвергается более быстрому флип-флопу, чем фосфолипиды. Следует отметить, что не только флип-флоп запускает функционально важные события в мембране. Латеральная и вращательная диффузия липидов оказывает регулирующее влияние на активность мембранных белков.
4. ФАЗОВЫЕ ПЕРЕХОДЫ ЛИПИДОВ В МЕМБРАНЕ
На все типы молекулярных движений липидных молекул сильное влияние оказывает структура, в которой в данный момент находится липвды бислоя тАУ гелеобразная или жидко-кристаллическая.
Липиды обладают замечательным свойством тАУ способностью к фазовым переходам в физиологических условиях. При определенных температурах, строго характерных для каждого вида липидов, липидные мицеллы могут быть в ВлтвердомВ» кристаллическом, организованном, гелеобразном состоянии или в ВлжидкомВ», мезофазном, так называемом жидко-кристаллическом состоянии. Жидкие кристаллы тАУ это анизотропные жидкости, так как оптически они сходны с кристаллами, проявляя разные свойства в разных направлениях, а механически сходны с жидкостью, они текут в зависимости от вязкости.
От состояния липидов в мембране зависит уровень молекулярной организации. Липиды в кристаллическом состоянии могут быть упакованы в кубический или гексагональный кристалл. Жидко-кристаллическая организация липидов в мембране очень разнообразна тАУ это так называемые нематики, смек-тики, холестерики.
Нематики тАУ наименее упорядоченная организация жидко-кристаллического состояния липидов. Молекулы нематика при умеренной температуре стремятся ориентироваться вдоль одного направления. В нематике очень многие молекулы одинаково ориентированы, их продольные оси параллельны друг другу, но такие области существуют недолго и границы их размыты. Области с одинаковой ориентацией молекул непрерывно рождаются и исчезают, что зависит от многих условий тАУ внешних границ, включений и различных воздействий. Магнитное и электрическое поля ориентируют молекулы нематика, причем выстраивают молекулярные оси параллельно своему направлению.
Смектики тАУ похожи на мыльные пленки, они более организованы, чем нематики, их молекулы образуют слои. В каждом индивидуальном слое молекулы передвигаются вдоль плоскости, все плоскости слоев находятся на одном и том же расстоянии. Смектики очень пластичны. Так, смектик в нативной мембране при охлаждении превращается в нематик.
Спиральная упаковка молекул вносит новое в ориентацию оптической оси жидкого кристалла. У холестериков тАУ слоистое строение с различным шагом спирали. Холестерическую спираль обозначают нередко как твист-ориентацию. Разбавление холестерика и увеличение шага спирали приводит к нема-тику. Оптическая активность холестериков очень велика, они избирательно отражают свет в зависимости от температуры, механической нагрузки, примесей, электромагнитных полей.
Жидкие кристаллы, сочетая в себе упорядоченность твердого тела и подвижность жидкости, отличаются высокой чувствительностью к внешним воздействиям, температуре, примесям, свету, внешним полям, они очень пластичны и очень долго хранят информацию. Эти свойства приобретают первостепенное значение в мембранах нервной ткани, где изменения электрических свойств лежат в основе проведения возбуждения.
Фазовый переход липидов является эндотермическим процессом, сопровождающимся изменением энтропии и энтальпии. Липидным структурам присущ лиотропный мезоморфизм и термотропный мезоморфизм. Оба свойства связаны между собой. Фазовый переход липидов Влгель тАУ жидкий кристаллВ» осуществляется при температуре, значение которой зависит от содержания воды в системе. Оно минимально, если общее содержание воды превышает то количество, которое могут связать липидные структуры. В то же время при температуре выше критической липиды могут находиться в упорядоченном состоянии при недостатке воды. Перекисное окисление липидов, увеличивающее содержание воды в бислое, существенно влияет на фазовое состояние мембраны.
Термотропные фазовые переходы липидов в мембране происходят в сравнительно широком температурном интервале. Это обусловлено тем, что в бислое одна фаза обязательно возникает в матриксе другой. Сосуществование в липидном бислое двух фаз устанавливает между ними сложное равновесие, приводя к снижению степени кооперативноетДв перехода. Обычно кооперативные фазовые переходы липидов в мембране затрагивают несколько сотен молекул. В нативной мембране постоянно находится большое число кооперативных единиц той или иной фазы. Этот полиморфизм является мощным регулятором транспортных систем мембраны.
Следует отметить, что на температуру фазового перехода большое влияние оказывают структура липидной молекулы, длина углеводородного скелета, наличие цис- и транс-двойных связей, структура полярных групп.
При переходе в жидко-кристаллическое состояние имеет место несколько одновременных событий: возрастает подвижность полярных групп липидов, увеличивается вращательная подвижность жирнокислотных радикалов относительно СтАУС-связей, увеличивается скорость латеральной диффузии. Это приводит к изменению геометрических размеров бислоя из-за латерального расширения площади, занимаемой каждой молекулой липида. Например, площадь, занимаемая 2С]6-фосфати-дилхолином, меняется от 0,49 до 0,58 нм, среднее расстояние между цепями увеличивается от 0,49 до 0,52 нм, а толщина углеводородного скелета уменьшается почти на 0,5 нм, т.е. латеральное расширение компенсируется утончением слоя. Гидрофобный объем мембраны увеличивается примерно на 1,5%.
В результате этих и ряда других изменений состояния липидов в мембране создаются особые условия для проникновения гидрофобных вешеств, изменения работы ионных каналов, внедрения в мембрану различных белков.
Микрогетерогенность бислоя и образование в нем кластеров молекул липидов способствует проявлению такого явления, как разделение фаз в мембране. Латеральное разделение липидных молекул в плоскости бислоя тАУ важная особенность мембраны. Особая сегрегирующая роль в мембране принадлежит холестерину. При низких концентрациях его в мембране происходит латеральное разделение фосфолипид-холе-стсриновых комплексов и свободных молекул фосфолипидов. При этом холестерин взаимодействует в первую очередь с теми молекулами фосфолипидов, которые имеют низкую температуру фазового перехода. Благодаря этому в бислое будут существовать области только жидкие и только твердые, а также области, где обе фазы сосуществуют. Наличие таких жидких и твердых областей в пределах мембраны изменяет ее сжимаемость, что сказывается на глубине погружения мембранных белков и на эффективности работы мембранных насосов.
Необходимо отметить, что кроме сегрегирующего холестерин проявляет и другое важное влияние на структуру и физические свойства липидного бислоя. Встраивание холестерина в фосфолипидный бислой вызывает как нарушение квазикристаллической упаковки цепей, так и уменьшение подвижности цепей тАУ Эти эффекты холестерина называют, соответственно, ВлразжижающимВ» и ВлконденсирующимВ». При температуре, превышающей точку фазового перехода фосфолипида, холестерин уменьшает подвижность углеводородных цепей. При добавлении холестерина площадь молекулы лецитина уменьшается с 0,96 до 0,56 нм. Вот почему высокое содержание холестерина характерно для миелина и плазматических мембран, тогда как внутриклеточные мембраны содержат его в небольших количествах. В плотных миелиновых мембранах фосфолипиды и холестерин содержатся в отношении 1:1, а в менее плотных мито-хондриальных мембранах это отношение равно 3:1 или 8:1. Этот уплотняющий эффект холестерина максимален в районе центрального участка жирнокислотных радикалов и ослабевает в направлении концевых метальных групп. При температуре ниже точки фазового перехода фосфолипидов холестерин разжижает углеводородную область бислоя.
Фазовые переходы липидов при постоянной температуре могут быть вызваны изменениями заряда полярных групп липидов, возникающими при изменениях рН, ионной силы, концентрации ионов. Доказано, что температура фазового перехода есть функция величина заряда и плотности заряда на липидной молекуле. Любое увеличение заряда полярных групп благоприятствует жидкому состоянию из-за латерального электростатического отталкивания, тогда как уменьшение заряда обусловливает переход в твердокристаллическое состояние.
Важным путем изменения поверхностного заряда липидов в физиологических условиях является адсорбция катионов. Связывание катионов заряженными липидами сильно зависит от поверхностного потенциала, значительно различающегося в твердом и жидком состояниях из-за различий в молекулярной упаковке.
Освобождение или адсорбция катионов на мембранной поверхности может запускать фазовые переходы липидов. При определенных физиологических условиях структурные изменения липидов могут вызывать освобождение двухвалентных катионов с поверхности мембраны. Так, при переходе гель тАУ жидкий кристалл с липидной поверхности освобождаются ионы кальция. Са+ 'и Mg+ стабилизируют организованную структуру, увеличивая температуру фазового перехода, а одновалентные катионы оказывают противоположный эффект. Двухвалентные катионы благоприятствуют гелеобразному, а одновалентные тАУ жидкому состоянию мембраны. Поверхность липидов может рассматриваться как резервуар катионов, который способен регулироваться структурными изменениями.
■ Подводя итог вышеизложенному, можно заключить, что в организации липидов, в их асимметричном размещении, подвижности, модификации внутримолекулярных взаимодействий сокрыты многообразные регулирующие возможности.
5. РОЛЬ БЕЛКОВ В ДИНАМИЧНОСТИ ЛИПИДНОГО БИСЛОЯ
Рассматривать динамичность бислоя мембраны без связи с белками нельзя. При липидных структурных перестройках в процесс вовлекаются интегральные, периферические и поверхностные белки мембраны. Более того, белки могут выступать в роли триггеров температурных структурных перестроек мембран, и белку часто принадлежит ведущая роль не только в инициации, но и в реализации структурной перестройки.
Одна из функций липидов в мембране тАУ придание белкам через межмолекулярные взаимодействия оптимальной конформации для функциональной активности. Липиды могут непосредственно участвовать в катализе. Липидный бислой определяет размещение белков, создает условия для их латерального перемещения и через фазовые переходы выполняет регуляторные функции. Жидкостность липидов влияет как на вращательную, так и диффузную свободу интегральных белков и их способность подвергаться конформационным изменениям. Вращательная и латеральная диффузия белков является отчасти следствием латерального движения мембранных липидов. Широкий спектр липидных молекул делает возможным широкое разнообразие специфических взаимодействий с мембранными белками.
Внедрение белка в фосфолипидный бислой упорядочивает его тАУ в результате структура бислоя становится более жесткой.
Считается, что это происходит за счет прилипания и ориентации фосфолипидных молекул, примыкающих к поверхности белка, ограничивающего подвижность этого слоя. У многих мембранных белков те их части, которые погружены в липид-ный бислой, особенно богаты гидрофобными аминокислотами, что повышает устойчивость их связей с липидами и фиксирует их ориентацию в мембране.
В бимолекулярном слое имеется два пула липидов, подвергающихся существенно различным скоростям диффузии. Один пул липидов находится в короткорадиусном взаимодействии с белками и потому подвергается ограниченной латеральной диффузии. Короткорадиусные взаимодействия могут быть очень специфичными и их может осуществлять только определенный тип липидов с особыми белками. Такие специфические липиды необходимы, в частности, для активации мембранных ферментов; они выступают здесь в качестве аллосте-рических эффекторов. Так, белковую молекулу Na+, К+-АТФа-зы окружает кольцевой слой липидов из 30тАУ32 молекул. Применение разнообразных физических методов показало, что кольцевые липиды могут многократно обмениваться с общим липидным пулом мембраны. Время обмена таких прочно связанных липидов с соседними молекулами составляет 10~ тАУ 10~ с. Это несоизмеримо меньше продолжительности одного ферментного цикла. Кроме того, оказалось, что сама фракция кольцевых липидов очень гетерогенна по своей обме-ниваемости, по фазовому состоянию и по способности к реактивации белка. Как минимум, роль кольцевых липидов заключается в поддержании строго определенного гидрофобного окружения данного белка.
В области температурных фазовых переходов таких липидов отмечается изменение каталитических и транспортных свойств белков. Общая доля кольцевых липидов довольно велика тАУ около 20%. Доказано, что можно изменять активность мембранных белков изменением связанных с ними липидов.
Другой пул липидов, удаленных от белков и подвергающихся быстрой латеральной диффузии, характерной для билипидного слоя, не пронизанного белком, составляет около 80%. Действие этих липидов на мембранные белки аналогично растворяющему эффекту воды на свойства растворимого белка.
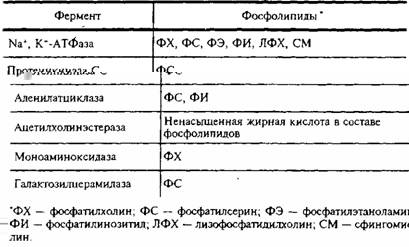
Приведем примеры функциональной роли индивидуальных липидов в мембранах ЦНС. В табл. 5 представлены данные о влиянии различных фосфолипидов на активность мембранных ферментов.
Активирование отдельными фосфолипидами мембранных ферментов
6. УЧАСТИЕ ЛИПИДОВ В РЕЦЕПЦИИ И ПЕРЕДАЧЕ ВНУТРЬ КЛЕТКИ СИГНАЛА
Межклеточные контакты, без которых немыслима деятельность ЦНС, обеспечивают постоянную передачу информации через плазматическую мембрану. Эта передача не может не касаться билипидного слоя. Процесс передачи информации через мембрану включает рецепцию внешнего химического сигнала и трансформацию его во внутриклеточный эффект.
Возникает вопрос, принимают ли участие липиды бислоя в рецепции внешних сигналов. В последнее десятилетие установлено, что сульфоиереброзиды играют довольно специфическую роль в рецепции опиоидов. Частично очищенный препарат рецептора опиоидов содержит высокую концентрацию сульфатидов. Предполагают, что сульфатная группа це-реброзидсульфата входит в состав или соседствует с активным центром опиатных рецепторов, который имеет белковую природу. Возможно, что взаимодействие опиатов с цереброзидсульфатами выполняет вспомогательную функцию, способствуя сосредоточению лигандов в области центра белковой природы.
При исследовании ряда кислых липидов только сульфатиды проявляли наивысшее сродство к опиатам в различных физиологических условиях. Доказательством важной роли сульфати-дов в рецепции опиоидов может служить и тот факт, что антитела к цереброзидсульфату, введенные в мозг крысы, снимали наркотическое действие морфина.
Если липиды бислоя могут быть участниками процесса рецепции, то естественно ожидать их участия в каскаде реакций, возникающих после активации рецепторов. М.Н. Хокин и Л. Э. Хокин впервые связали холинергическую стимуляцию с усилением обмена фосфатидилинозита и фосфатидной кислоты. Явление получило название Влфосфолипидного эффектаВ»; этот термин сейчас заменен на термин Влфосфоинозитид-ный эффектВ», поскольку появилось большое число работ, показывающих именно их регуляторную роль в транспорте вторичного мессенджера тАУ ионов кальция тАУ через мембраны.
Содержание фосфоинозитидов в мембранах ЦНС не превышает 0,5тАУ2% от общих липидов, локализованы они преимущественно в плазматических мембранах, в миелине, в эндоплазматическом ретикулуме, наружной митохондриальной и ядерной мембранах. В состав фосфоинозитидов входит арахидоновая кислота, являющаяся важным источником простагландинов. Деполяризация мембраны приводит к быстрому высвобождению арахидо-новой кислоты именно из фосфоинозитидов. Р. Митчелл высказал гипотезу о прямой связи расщепления фосфоинозитидов с рецепторным аппаратом клетки и увеличением внутриклеточной активности. В синаптосомах стимуляция части мускариновых и ах-адренерги-ческих рецепторов обусловливает фосфоинозитидный эффект, сопровождающийся изменением проницаемости плазматической мембраны для ионов кальция.
Участие фосфоинозитидов и продуктов их обмена в регуляции транспорта кальция осуществляется несколькими путями:
1) при распаде фосфатидилинозитидов образуется 1,2 тАУ диа-цилглицерин, стимулирующий активность протеинкиназы С, которая, в свою очередь, фосфорилирует белок Са-каналов и некоторые другие белки;
2) трифосфоинозитол, освобождающийся при расщеплении фосфатидилинозитидов, обладает высокой способностью связывать двухвалентные катионы; по этой причине он индуцирует мобилизацию мембранно-связанного кальция;
3) инозитол-трифосфат способен также повышать уровень внутриклеточного кальция за счет открытия кальциевых каналов эндоплазматического ретикулума. Таким образом, происходит сопряжение выброса кальция из внутриклеточных мест хранения с входом кальция через мембраны.
До включения описанного механизма концентрация свободного кальция в цитоплазме нейрона составляет примерно 1 тАУ .10~М. Концентрация кальция снаружи нейрона в десятки тысяч раз выше. Мобилизация Са+ из внутриклеточных и внеклеточных источников в сотни-тысячи раз повышает его уровень в цитоплазме. Повышенный уровень Са+ служит активатором ряда процессов, в том числе некоторых протеинкиназ.
На молекулярном уровне передача этого сигнала через мембрану осуществляется цепочкой мембранных белков, последовательно взаимодействующих друг с другом для передачи сигнала малым молекулам, находящимся в цитоплазме. Информация от рецептора на поверхности клетки передается так называемому G-белку, который активирует фермент фосфодиэстеразу, расщепляющую трифосфоинозитид до инозитол тАУ 1,4, 5-трифосфата и 1,2 тАУ диацилглицерина. Инозитолтрифосфат растворим в воде, диффундирует в цитоплазму, где и вызывает описанное выше освобождение кальция. Освободившийся кальций участвует в активации протеинкиназ.
Липофильный диацилглицерин, отличный по своему жирно-кислотному составу от стабильного пула диацилглицеринов, остается в мембране, изменяет ее текучесть и, как уже упоминалось, активирует мембранно-связанную протеинкиназу С,
Эти две различные ветви фосфоинозитидного цикла ведут в конечном счете к фосфорилированию двух различных наборов белков. Оказалось, что с помощью активирующих веществ каждую из ветвей цикла можно привести в действие независимо друг от друга. С другой стороны, применение сочетанного действия фор-боловых эфиров и кальциевых ионофоров помогло установить синергизм двух сигнальных ветвей инозитидного цикла. В таком раздвоенном сигнальном пути совместным действием веществ можно запустить большое число внутриклеточных процессов.
В дальнейшем образовавшиеся 1,2 тАУ диапилглицерин и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается.
7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин тАУ уникальное образование, организация которого позволяет проводить импульс в аксоне с минимальной затратой энергии. Миелиновая оболочка тАУ высокоорганизованная многослойная структура, состоящая из сильно растянутой и модифицированной плазматической мембраны олигодендроглиальной клетки.
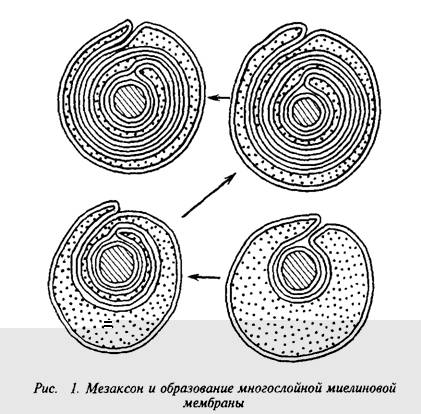
Плазматическая мембрана олигодендроцита образует вокруг аксона сложную мембранную структуру тАУ мезаксон, который является элементарной единицей миелина, имеет пятислойную структуру: белок-липид-белок-липид-белок. Эта пя-тислойная структура, многократно закручиваясь вокруг аксона,
конденсируется в компакт
Вместе с этим смотрят:
G-белки и их функция
Австралопитеки - обезьянолюди или человекообезьяны?
Адаптация микроорганизмов в экстремальных условиях космоса
Адвентивна флора Чернiгiвськоi областi: iсторiя формування та сучасний стан
Адсорбция ионных и неионных поверхностно-активных веществ (ПАВ)