Ферменты дереворазрушающих грибов
Структурный каркас почти всех наземных растений состоит из полимеров: лигнина, гемицеллюлозы, целлюлозы. Объединяясь в различных пропорциях, они образуют лигноцеллюлозный материал. На его долю приходится основная часть биомассы, остающейся в огромных количестве в виде отходов сельского хозяйства, деревообрабатывающей промышленности и других отраслей хозяйственной деятельности человека (древесина, солома, рисовая шелуха, использованная бумага, картон и т.д.). Эти отходы необходимо перерабатывать или использовать в качестве промышленного сырья [1].
Грибы всегда вызывали интерес человека загадочностью своей природы. Их плодовые тела несущие миллиарды спор, появляются на почве, на деревьях, на травянистых растениях, включая культуры [2]. Они относятся к третьему царству природы Fungi или Mucota. Их относят к главным мусорщикам тАУ утилизаторам мертвого органического вещества [3].
Превращение древесины в природе сводится в конечном итоге к ее полному разложению и гумификации. Основную роль в нём безусловно играют различные грибы-ксилотрофы. На их долю приходится более 90% разлагаемой древесины. Несмотря на то, что бактерии обладают широким набором целлюлаз, гемицеллюлаз и пектиназ, они в очень ограниченной степени разлагают лигнин [4].
Благодаря мощной внеклеточной ферментативной системе грибов-ксилотрофов позволяют им утилизировать труднодеградируемые полимеры клеточных стенок древесины вплоть до полного разложения. Это стимулирует интерес к их интенсивным исследованиям [5]. Традиционно ксилотрофы рассматриваются как источники разнообразных ферментов для трансформации древесины. В настоящее время перспективно использовать как транспортные средства, способные доставить ферменты в нужные компартменты древесной клетки. Это может использовать для направленной трансформации древесины, получения смешанных композитов высокой прочности. В качестве потенциальных связующих в древесных композитах, за счет образуемых ими красителей хиноидной структуры, склонные к дальнейшей конденсации [3].
Выращивание ксилотрофов на лигноцеллюлозных отходах можно получать не только дополнительный источник белка, но и ценные компоненты для медицины, получаемые из плодовых тел, мицелия (полисахариды, стерины). При этом решается проблема загрязнения окружающей среды.
В связи с этим были рассмотрены основные представители ксилотрофных базидиальных грибов, их лигнолитический и целлюлолитический ферментативные комплексы, и их применение в промышленности.
1.Характеристика дереворазрушающих грибов
1.1Грибы, участвующие в разрушении древесины
На древесину в процессе эксплуатации воздействует целый ряд факторов окружающей среды, приводя к ее старению и разрушению. Среди них: климатические (УФ тАУ излучение, влажность, ветровые нагрузки, кислород воздуха) и биологические (грибные поражения, поражения насекомыми, бактериями, водорослями). Процесс деструкции заложен самой природой для поддержания экологического равновесия, поэтому в естественных условиях древесина, с течением времени, разрушается до углекислого газа и воды тАУ самых простых химических соединений [6]. Грибы приспособлены к существованию в порах плотного субстрата, бедного питательными веществами, но содержащие трудноусваиваемые источники углерода тАУ лигнин и кристаллическая целлюлоза [4].
Грибы, развивающиеся на древесине (ксилофилы, ксилотрофы), практически все принадлежат к трем классам высших грибов, имеющие разделенные на клетки (септированные) гифы. Это аскомицеты (Ascomycetes, сумчатые грибы), дейтеромицеты или несовершенные грибы (Deuteromycetes, Fungiimperfecti), и базидиомицеты (Basidiomycetes) тАУ наиболее сильные разрушители [2].
На первой стадии при поражении, на древесине появляются грибы, питающиеся соками живого дерева. Такие как плесневые грибы Penicillium, Aspergillus, живущие на поверхности древесины. Затем в подготовленных плесневыми грибами оптимальных условиях начинают размножаться деревоокрашивающие грибы. Завершают процесс разложения древесины дереворазрушающие грибы. Они вызывают сильное гниение древесины, приводящее к появлению продольных и поперечных трещин, а затем и минерализации древесины [7].
У плесневых грибов поверхностная часть грибницы развивается на поверхности древесины и образует на ней налет в виде скопления окрашенных спор, мицелия и органов спороношения, как показано на рисунке 1.1. На древесине обычно встречается плесень зеленоватая и белая, но иногда розовая, желтая или темная. Плесневые грибы развиваются в температурном диапазоне 24В°С − 30В°С. Они являются возбудителями окислительного брожения. В качестве промежуточных продуктов этого биохимического процесса образуются органические кислоты, такие как глюконовая, фумаровая, винная, яблочная, щавелевая, янтарная и лимонная. Эти кислоты разъедают органические материалы, т.е. древесину. Основные виды плесневых грибов: Sporotrichum, Trichoderma, Penicillium, Mucor, Thamnidiu, Cladosporium.
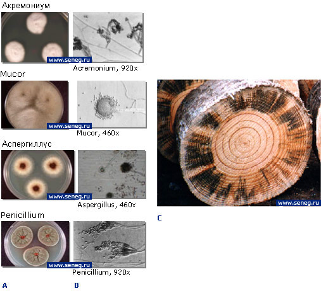
Рисунок 1.1 − Плесневые грибы:
A) Вид колонии на чашке; B) Под микроскопом; C) На древесине
Деревоокрашивающие грибы вызывают специфическую синевато-серую окраску заболони, называемую ВлсиневойВ», в соответствии с рисунком 1.2.

Рисунок 1.2 − Деревоокрашивающие грибы
А) Колонии; B) Под микроскопом; C) На древесине
Макроскопические признаки поражения древесины этими грибами в виде окраски обычно проявляются уже на 2-3 сутки после инфицирования. Это обусловлено тем, что молодой мицелий бесцветен и начинает выделять типичный пигмент не сразу. На поверхности древесины может развиваться воздушный мицелий и органы спороношения в виде пушистого или порошкообразного окрашенного налета [6].
В зависимости от характера заражения и распространения в древесине гиф грибов различают поверхностную и глубокую синеву. Ограниченное распространение грибов в глубь древесины связано с задержкой их роста в результате подсыхания древесины. Глубокие окрасы проникают в древесину более чем на 2 мм. Среди них различают сплошные, охватывающие всю заболонь (глубокая синева) и пятнистые, поражающие отдельные участки заболони. Глубина залегания окраски при подслойной синеве зависит от вида гриба, размера свойственной ему зоны бесцветного молодого мицелия (зоны скрытой синевы), ширина которой колеблется от 5 до 12 мм. Грибы-возбудители синевы, попавшие из воздуха на поверхность свежеспиленной древесины в виде спор, не дают сразу окраски в течение двух и более недель (период бесцветной, скрытой синевы). Окрашивание появляется на третий-четвертый день. Деревоокрашивающие грибы оптимально развиваются в диапазоне влажности 50-90%. В древесине, насыщенной водой, деревоокрашивающие грибы не способны развиваться из-за отсутствия кислорода. Для прорастания грибов этой группы необходима высокая влажность и аэрация [4].
Основными возбудителями синевы на хвойных породах являются грибы из класса Ascomycetes: Ophistomacoerulea, O. piceae, O. pini, Endoconidiophora. и из класса Deuteromycetes: Hormonemadematiodes, Trichosporiumtingens, Claosporiumherbarum, а также грибы следующих групп: Stemphulium, Cladosporium, Alternaria, Sporodesmium, Phialophora, Aposhaeria, Discula, Burgoa, Leptographium, Sortaria, Verticillium, Fusarium, Aspergillius, Penicillium, Paecilomyces, Trichoderma, Chaetomium, Trichosporium, Pullularia [9]. Эти грибы вызывают разрушения древесины по типу Влумеренной гнилиВ», вызывая разрушения анатомически различного характера, в разной степени снижают механические свойства древесины. Глубина поражения этими грибами составляет 0,5-3 мм. Особые деструктирующие гифы, в соответствии с рисунком 1.3, способны поражать стенки паренхимных клеток сердцевинных лучей и смоляных ходов, что приводит к увеличению скорости водо- и влагопоглощения древесины [6].

Рисунок 1.3 − Плесневые и деревоокрашивающие грибы под
электронным микроскопом
Деревоокрашивающие грибы в различной степени способны изменять свойства древесины. При дальнейшем развитии грибов многие из них способны разрушать стенки сердцевидных лучей и вторичных слоёв клеточных стенок по типу, близкому к умеренной гнили [9]. Деревоокрашивающие грибы -это только начало процесса. Они способствует дальнейшему разрушению древесины дереворазрушающими грибами [6].
Некоторые классы грибов могут разрушать клеточные стенки древесины и существенно изменять ее физико-механические свойства. Такой процесс называется гниением древесины, а вызывающие его грибы тАУ дереворазрушающими, пример на рисунке 1.4. Гниение является основной причиной разрушения древесины [10].
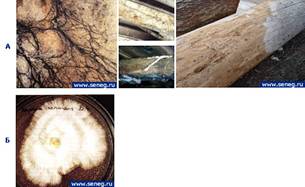
Рисунок 1.4 − Дереворазрушающие грибы на древесине
А) Вид поврежденной древесины; Б) Колонии Serpula lacrimans
Возбудители деструктивных гнилей, разлагающих целлюлозу, проникают в клетки древесины из сердцевинных лучей отдельными разветвленными гифами, а лигнинразрущающие грибы тАУ пучком гиф, который затем обильно разветвляется. Затем такой гриб заполняет полость клетки густым переплетение гиф, причем мицелий иногда собирается в клубки, как у Serpulafrustulosum [3]. Существует множество дереворазрушающих грибов. Они различаются между собой по форме, строению и окраске грибницы, шнуров, плодовых тел и спор, а также по скорости и силе разрушения древесины [3].
1.2Базидиомицеты тАУ основные представители дереворазрушающих
грибов
Наиболее сильными разрушителями являются грибы, относящиеся к классу базидиомицетов. Ксилотрофные базидиомицеты тАУ это грибы, образующие крупные плодовые тела (карпофоры), спорообразующий слой которых называют гименофором [1]. Среди них выделяют ксилопаразитов, развивающиеся на живой древесине и кустарниках. Например губки Heterobasidionannosum, паразитирующая на соснах и елях, ложный трутовик Phellinusigniarius, паразитирующий на лиственных породах особенно часто на осинах и ивах. Большая часть этой группы относится к ксилосапротрофам, развивающиеся на мертвой древесине. Например трутовик настоящий Fomesfomentarius, опёнок осенний Armillariellamellea [2].
На поверхности древесины они помимо воздушного мицелия образуют и другие вегетативные мицелиальные структуры. Основная форма размножения тАУ базидиоспоры. Они формируются в результате полового процесс в спорогенных клетках тАУ базидиях, располагающихся в плодовых телах тАУ базидиомах. Базидиома является специализированным талломным образованием из функционально дифференцированных тканей мицелия [3].
Дереворазрушающие грибы способны увлажнять древесину в процессе освоения за счет воды, образующейся при разложении целлюлозы. Возбудители биоповреждений древесины, относятся в основном к следующим группам грибов: Coniophora, Tyromyces, Zentinus, Serpula, Gloeophyllum, Trametes, Pleurotus, Schizophyllum [9].
Характер гниения зависит от того, какими ферментами гриб воздействует на древесину, какие компоненты клеточных оболочек и в какой последовательности он разрушает. В начале деятельности дереворазрушающих грибов внешний вид древесины не изменяется, и присутствие грибных нитей в ней можно обнаружить только под микроскопом, в тонком срезе. В дальнейшем древесина изменяет свой естественный цвет, становится желтой или красноватой, а затем бурой и коричневой. Плотность и прочность древесины постепенно снижаются, она становится лёгкой, мягкой, теряет вязкость. Гниль такого типа называют деструктивной [4].
Некоторые грибы, поражающие растущие деревья, вызывают гниль другого типа тАУ коррозионную, при которой вначале появляются небольшие светлые пятна и ямки, а затем древесина расщепляется на отдельные волокна. Эта группа грибов использует в первую очередь лигнин древесины, оставляя нетронутой целлюлозу, белые пятна и выцветы которой видны на поверхности среза. К коррозионной гнили также относятся сердцевинные гнили: пестрая, ямчатая, ситовая [6].
При развитии умеренной гнили поверхностные слои древесины теряют свою структуру и превращаются в мягкую темную грязеподобную массу. После подсушивания древесины в пораженном слое наблюдается сильное усыхание и появление мелких продольных и поперечных трещин. Все без исключения дереворазрушающие грибы вырабатывают фермент целлюлазу, расщепляющую целлюлозу [11]. В зависимости то состава ферментного комплекса условно подразделяют их на следующие: Белая гниль разрушает все структурные компоненты древесины, приводя к появлению характерного волокнообразного и бледного внешнего вида. Бурая гниль ВлраскалываетВ» целлюлозу, что вызывает расщепление древесины. Участок дерева, пораженный такой гнилью, становится коричневым. Дерево темнеет, трескается и рассыпается. Их представлено значительно меньшим числом видов, чем грибы белой гнили. Они преобладают в хвойных лесах и гораздо быстрее разрушают древесину [6].
Мягкая гниль. Гниение здесь в основном затрагивает древесину, контактирующую с почвой и находящуюся в морской среде. Сильнее всего поражается древесина с высоким содержанием влаги [6].
Возбудители белой гнили представлены базидиомицетами (афиллофоровым и пластинчатым), сохраняют форму и волокнистое строение древесины, но вызывают ее осветление вплоть до светло-желтого или белой окраски. Деградация сопровождается уменьшением прочности и увеличением набухания. К ним относят, например, сосновая губка. Большинство из них предпочитает лиственную древесину [3].
Возбудители бурой гнили относятся к базидиомицетам. Древесина под их действием буреет тАУ накапливаются ароматические хромофоры в видимой области спектра (хиноны, хинонметиды, карбоксильные группы, устойчивые феноксильные радикалы, переходит в видимую область спектра поглощение лейкохромофорных групп). Одновременно она становится хрупкой. Деструкция сопровождается продольным сморщиванием и деформацией клеточных стенок. Типичными возбудителями являются большинство домовых грибов, трутовики Швейница, серножелтый и окаймленный, лиственничная губка [9].
Возбудители мягкой гомогенной гнили разлагают в клеточной стенке древесины преимущественно вторичный слой с образованием темноокрашенных гумусовых веществ (шоколадного цвета), постепенно разрушая ее внутреннюю структуру, из-за чего влажная древесина приобретает консистенцию густой пасты легко царапаемой ногтем. Мягкую гниль вызывают некоторые аскомицеты несовершенные грибы. Они разрушают хвойную и лиственную древесину [3].
Грибы белой гнили известны также как активные деструкторы широкого ряда поллютантов, включая полихлорированные фенолы, нитро- и аминозамещенные фенолы, диоксины [12]. Это единственные эукариоты способные разлагать полициклические ароматические углеводороды (ПАУ). Например, они могут метаболизировать фенантрен, нафталин, антрацен, флюорантен, флюорен, пирен, бензaпирен, бензaантрацен, хризен, карбазол и многие другие [13]. В отличие от бактерий, они могут метаболизировать ПАУ с 4 и более конденсированными кольцами. В настоящее время неизвестно ПАУ полностью устойчивых к деградации грибами белой гнили [8].
Эти грибы не используют ПАУ, также как и лигнин, в качестве единственного источника углерода и энергии, но способны кометаболизировать их. Полная минерализация ПАУ грибами белой гнили ограничена и составляет, по данным разных авторов, от 0,17% до 10%, большая часть этих веществ метаболизируется в гидрофильные гидроксилированные продукты, например дигидродиолы, хиноны и 2,2'-дифеновую кислоту тАУ продукт расщепления ароматического кольца. Процесс деградации ПАУ, также как и процесс деградации лигнина является окислительным и протекает наиболее интенсивно при рН 4,0-5,0 [13].
Современные исследования деградации ПАУ грибами белой гнили направлены на детальное изучение метаболических путей и вовлеченных в них ферментных систем. Деградативную активность этих грибов в основном связывают с их внеклеточной лигнинолитической ферментной системой [11].
Для прикрепления к твердому субстрату и сохранения внеклеточных ферментов вблизи поверхности гифов грибы используют особые структуры тАУ гифальные чехлы, построенные из слизистых полисахаридов предположительно β-1,3-β-1,6-глюканов [4].
2. Лигнолитический ферментный комплекс дереворазрушающих
грибов
Способность грибов разрушать лигниновый компонент древесины известна давно. Наиболее активными деструкторами лигнина в природе являются грибы белой гнили, которые образуют гетерогенную группу, состоящую большей частью из базидиомицетов, относящихся к семействам Agaricaceae, Corticaceae, Pleurotaceae, Strophariaceae и нескольких аскомицетов [6]. Так как в качестве лигнинолитиков изучены преимущественно базидиомицеты, то в узком смысле под грибами белой гнили обычно понимают лигнинолитические базидиомицеты. Наиболее подробно изучены Phanerochaete chrysosporium, Coriolus versicolor и Pleurotus ostreatus. [4]
Хотя лигнин является потенциально богатым энергией материалом, он не может служить единственным источником углерода и энергии, его деградация возможна при наличии в среде ростового субстрата, такого как целлюлоза или глюкоза. Деградация лигнина является событием вторичного метаболизма и происходит когда в среде культивирования исчерпываются источники углерода, азота или серы. [14] Разложение лигнина тАУ процесс окислительный, стимулирующее действие кислорода на биодеградацию лигнинового полимера характерно для всех грибов белой гнили. Лигнинолитический процесс у этих грибов подвержен влиянию рН, максимум лигнинолитической активности приходится на рН 4,5 [13].
Структура высокомолекулярной полимерной гидрофобной молекулы лигнина определяет свойства расщепляющей его ферментативной системы. Такая система должна быть внеклеточной негидролитической и неспецифической. Лигнинолитическая система грибов белой гнили является внеклеточной окислительной и неспецифической, и включает лигнин пероксидазу, Mn-пероксидазу и лакказу [4].
Лигнин тАУ глобулярный нестереорегулярный нерастворимый полимер. Его молекулярная масса более 10 000. Состоит из остатков фенилпропана, соединенных несколькими типами углерод тАУ углеродных и эфирных связей. Молекулы этого ароматического вещества при образовании лигнина соединяются друг с другом случайным образом с помощью разных химических связей не подающиеся химическому расщеплению. У растений лигнин образует комплекс с гемицеллюлозой, в которой заключены проводящие пучки. Лигнин обусловливает ригидность растений, а также их устойчивость к механическим повреждениям и действию микробов [1].
С развитием биотехнологии базидиомицеты становятся доступным источником ферментов различных классов. Способность базидиомицетов синтезировать внеклеточные ферменты, характеризующиеся высокой активностью и стабильностью, дает возможность использовать их ферментные комплексы для практических целей [14].
В настоящее время во всем мире ведется интенсивная разработка технологий на основе базидиальных лигнинолитических грибов и их ферментов как для обработки лигниноцеллюлозных материалов, так и для утилизации лигнинсодержащих отходов, накапливающихся в природе в огромных количествах [15].
Дереворазрушающие базидиомицеты отличаются высоким содержанием окислительно-восстановительных ферментов, в первую очередь пероксидазы, Мп-пероксидазы, тирозиназы и лакказы [8]. Высшие базидиомицеты по составу лигнолитических ферментов объединяют в следующие группы:
В первую группу вошли грибы, обладающие лакказой, лигнин- и марганецпераксидазой активностью Phellinus рiпi, Trametes hirsuta, Bjerkandera adusta, Phanerochaete chrysosporium.
Вторая группа представлена грибами Lentinu edodes, Paus tigrinus, P. chrysosporium, Dichomitus squalens, обладающих Mn-пероксидазной и лакказной активностью.
Третья группа характеризуется лигнинпероксидазной и лакказной активностью тАУ грибы Trametes versicolor, Phlebia radiate, Pleurotus ostreatus.
Грибы, составляющие четвертую группу определены как Pleurotus ostreatus, P. eringii, В. adusta и для них характерна лакказа, арилалкогольоксидаза и другие ароматические оксидазы [12].
Разнообразное сочетание ферментативных комплексов у лигнинразрушающих грибов связано в первую очередь с экологическими особенностями грибов, трофической специализацией и является следствием длительной эволюции растений и грибов [7]. Представители различных таксономических и экологических групп обладают сходным составом ферментов. Однако уровень активности внеклеточных ферментов имеет существенную штаммовую и видовую вариабельность [10].
Внеклеточные лигнинолитические ферментные комплексы грибов белой гнили включают следующие типы окислительно-восстановительных ферментов: I. гем-содержащие ферменты, П. флавинсодержащие ферменты, III. целлобиозодегидрогеназа, IV. медьсодержащие ферменты [4].
Помимо лигнинразрушающих ферментов эти грибы образуют также системы гидролаз, гемицеллюлозы и целлюлозу [9].
I. Гемсодержащие ферменты,среди которых принято особо выделять лигнинпероксидазы (LiP) и марганец-пероксидазы (МnР). Основная функция этих ферментов тАУ прямое, как у LiP или опосредованное медиатором (вератролом у LiP и ионами Мn у МnР) одноэлектронное окисление ароматических субстратов до соответствующих радикалов и двухэлектронное восстановление перекиси водорода до воды [4].
Лигнинпероксидаза(диарилпропаноксигеназа, Н202-зависимая оксигеназа, лигниназа, КФ 1.11.1.14) тАУ гемсодержащий фермент с молекулярной массой 39-42 кДа, являющийся донором Н202. Катализирует различные реакции одноэлектронного окисления ароматических соединений (предпочтительно нефенольных) за счет кислорода перекиси водорода, с восстановлением ее до воды. К ним относят:
1. Сα−Сβ тАУ разрыв пропильной группы лигнина и модельных соединений.
2. Гидроксилирование бензильных метиленовых групп.
3. Окисление бензиловых спиртов до соответствующих альдегидов и кетонов.
4. Окисление фенолов и расщепление ароматического кольца в нефенольных модельных соединениях лигнина [16, 17].
Полученный арильный катион-радикал спонтанно разрушает различные связи в зависимости от структуры и наличия реактанов. Лигнинпероксидаза способна окислять и фенольные соединения. При этом образуются в результате окисления ферментов нефенольных соединений. Выделено около 15 изоформ данного фермента [16]. Лигнинпероксидаза обнаружена лишь у немногих базидиальных грибов: Phanerochaete chrysosporium, Trametes versicolor, Trametes hirsuta, Panus tigrinus, Coriolopsis occidentalis [13].
Мn-зависимая пероксидаза тАУ гемсодержащий фермент (КФ 1.11.1.3), участвующий в деполяризации синтетического лигнина [16]. Она окисляет фенольные соединения в присутствии перекиси водорода. Проявляет активность в среде, содержащей Мn, имеет молекулярную массу 46 кДа [8]. Впервые выделена из культуральной жидкости гриба Ph. chrysosporium. Принцип функционирования фермента заключается в окисление Мn2+ до Мn3+ с использованием пероксида водорода в качестве окислителя. Активность фермента стимулируется простыми органическими кислотами, которые стабилизируют ион Мn3+. В результате взаимодействия последнего с кислотами образуется хелат, обладающий окислительными свойствами [16]. Регенерация Мn осуществляется сопряженной реакцией разложения перекиси водорода. При отсутствии в среде Н202 Мn-зависимая пероксидаза способна продуцировать пероксид водорода. Мn-зависимая пероксидаза отмечена у таких грибов как Trametes versicolor, Phlebia radiate, Dischomitus squalens и др. [13].
П. Флавинсодержащие ферменты,осуществляющие, в основном двухэлектронное восстановление молекулы кислорода до перекиси водорода и одновременно двухэлектронное окисление ОН-группы, соответствующих субстратов, до карбонильных групп. Среди них тАУ глюкозооксидазы, пиранозо-2-оксидазы, метанол-оксидазы, арилалкогольоксидазы [13].
Глюкозооксидаза тАУ флавинсодержащая оксидаза с молекулярной массой 80 кДа, катализирующая окисление глюкозы, восстанавливая при этом кислород до перекиси водорода. Глюкозооксидаза выявлена у грибов Paus tigrinus, P. Chrysosporium [8].
Пираноза-2-оксидаза ароматических спиртов, катализирует окисление низших первичных и ароматических спиртов до соответствующих альдегидов и пероксида водорода [13].
Арилалкогольоксидаза(вератрил-алкоголь оксидаза), внеклеточная флавин-оксидаза с молекулярной массой 72,5-78 кДа, катализирует окисление ароматических спиртов до альдегидов, восстанавливая кислород до перекиси водорода. Арилалкогольоксидаза отмечена у грибов, вызывающих белую гниль древесины: Phanerochaete chrysosporium, Trametes versicolor [13].
Метанолоксидаза(МеО), внеклеточный флавин-содержащий фермент с молекулярной массой 75 кДа, катализирует окисление метанола, выделяющийся при деградации лигнина в реакциях деметилирования фенолов, до формальдегида, восстанавливая кислород до перекиси водорода. Выделена и охарактеризована у Phanerochaetechrysosporium, Trametesversicolor, Panustigrinus[13].
III. Медьсодержащие ферменты. Разделяют на две группы.
III.1 Голубые оксидазы тАУ лакказа и родственные или им неголубые оксидаза, осуществляющие четырехэлектронное восстановление кислорода до воды с параллельным одно- или двухэлектронным окислением преимущественно фенольным субстратом до соответсвующих феноксирадикалов или хинонов. [8]
Катехолоксидазаучаствует в гидроксилировании монофенолов до дифенолов и окислении дифенолов в ортохиноны. Она широко распространена среди микроорганизмов и имеет названия, соответствующие катализируемому субстрату: монофенолоксидаза, полифенолоксидаза, фенолаза, крезолаза, тирозиназа и т. п. [13].
Лакказы (бензидол: кислород оксидоредуктаза, КФ 1.10.3.2) тАУ голубая медьсодержащая оксидаза, которая катализирует четырехэлектронное окисление фенолов, ароматических аминов и фенилендиамина, используя кислород как акцептор электронов с восстановлением его до воды. Практически все исследованные лакказы тАУ это мономеры, гликопротеины с углеводной частью от 1 до 15% массы фермента, которая состоит из остатков маннозы и N-ацетилглюкозамина [16]. Окисляют широкий круг субстратов преимущественно фенольной природы с образованием феноксильных радикалов, а также нефенольные соединения в присутствии специфических электронных переносчиков ABTS или HBT (1-гидроксибензотриазол) [16]. Лакказы древоразрушающих грибов имеют более низким Ph-оптимумом окисления фенольных субстратов, совпадающим с оптимум действия большинства грибных целлюлаз. Так, для лакказ Gaeumannomycesgraminisvar. tritici, древоразрушающего базидиомицета PM1, Coriolushistris он близок к 4,5Ph, для лакказ других трутовиков (C.zonatus, Tramenassanguinea, Polyporuspinsitus) составляет 4,9 Ph -5Ph [4].
III.2 Медьсодержащие ферменты, осуществляющие двухэлектронное восстановление кислорода до перекиси с одновременным двухэлектроным окислением подходящей спиртовой или альдегидной группы (глиоксальоксидаза, галактооксидаза) [8].
Наибольшей оксидазной активность обладают штамма TrameteshirsuteD-5 и GerrenaunicolorМ-2 [18].
Лигнин под действием бурых гнилей преимущественно гидроксилируется и деметилируется и в меньшей степени депомилиризуется. Окисление непосредственно идет до углекислоты с изъятием углерода в основном из боковых цепей и метоксильных групп [13].
Предположительно, лигнинразрущающие ферменты действуют на поверхности гиф, находящихся в контакте с клеточной стенкой. Вместе с тем, разложение наблюдается не только в местах контакта, но и по всей поверхности люмена, если там находятся всего одна-две гифы гриба, и даже в толще вторичной оболочки. Снижение выхода низкомолекулярных продуктов окисления лигнина (ароматических кислот и альдегидов) после обработки древесины лигнинразрушающими грибами объясняет их окислительным и конденсирующим действием. Однако разные грибы в разной степени окисляют и конденсируют остаточный лигнин [4].
Так было показано, что выявленная на поверхности мицелия внеклеточная лакказа штамма гриба PleurotusostreatusDl могут участвовать в начальных этапах метаболизма полициклических ароматических углеводородов (в данной работе хризена). Тогда как гибридная Mn-пероксидаза окисляла образующиеся метаболиты (фталевая кислота) [12].
3. Целлюлитический комплекс дереворазрушающих грибов
Целлюлоза очень ценный материал из которого можно получать множество продуктов, например этанол. Она является наиболее простым компонентом лигноцеллюлозного материала и самым распространенным природным полимером. Его длинные цепи состоят из остатков D-глюкозы соединенных β-1,4 связями. Целлюлоза отличается по строению от крахмала. В целлюлозе полимерные цепи упакованы так, что образуют кристаллоподобную структуру, непроницаемую для воды. Поэтому она не растворяется в воде и устойчива к гидролизу [1].
Целлюлолитические ферменты, осуществляющие биодеградацию целлюлозы, самого распространенного биополимера на Земле, занимают центральное место в круговороте органического углерода [2]. Основными микроорганизмами, продуцирующими целлюлазы, являются грибы возбудители мягкой и бурой гнили, а также различные виды аэробных и анаэробных бактерий. История исследования целлюлаз насчитывает уже более 50 лет, В течение этого периода важнейшим свойством, характеризующим целлюлазный комплекс, считалась его способность к глубокой деструкции целлюлозосодержащих субстратов (так называемая ВлсахаролитическаяВ» активность). Поэтому исследования, в основном, были направлены на поиск ферментных препаратов и их продуцентов, эффективно осуществляющих гидролиз целлюлозы до глюкозы [19]. Целлюлазы этих препаратов, как правило, проявляли максимальную активность в кислой среде (рН 4-5) [14], но различались по субстратной специфичности, адсорбционной способности и термостабильности. Целлюлазы находят все более широкое применение в текстильной, целлюлозно-бумажной, пищевой и других отраслях промышленности [6]. В последнее время усилия исследователей направлены на поиск целлюлолитических ферментов, способных мягко воздействовать на поверхность целлюлозного субстрата, не приводя к глубокой деструкции целлюлозной матрицы [4]. Обнаружение ферментов с такой (тополитическая активность) активностью открыло новые возможности их применения. Например, для депигментации джинсовых изделий с целью придания им более привлекательных потребительских свойств (альтернатива традиционным химическим способам ВлваркиВ», а также обработке пемзой); для биополировки текстильных материалов с целью удаления микродефектов и ворса; как компонента моющих средств и т, д. В связи с этим наиболее перспективными для использования являются так называемые ВлнейтральныеВ» целлюлазы, демонстрирующие высокую активность [20].
Разрушение материала клеточных стенок под действием ферментов грибов наблюдается по появлению зон растворения вокруг гиф. Зона растворения постепенно увеличивается и гифа проникает сквозь внутренний слой клеточной стенки [4].
Всего выделяют 3 основных типа целлюлаз, которые катализируют различные типы реакции:
а) Эндоцеллюлаза разрывает внутренние связи, для того чтобы нарушить кристаллическую структуру целлюлозы. В результате образуются отдельные полисахаридные цепи целлюлозы.
б) Экзоцеллюлаза отщепляет от концов 2 или 4 остатка, в результате образуются тетрасахара или дисахариды, состоящие из целлобиозы. Выделяют два основных типа экзоцеллюлаз (или целлобиозодегидрогеназ). Первый тип отщепляет от редуцирующего конца. Второй тип от нередуцирующего конца целлюлозы.
в) Целлобиоза или бета-глюкозидаза гидролизуют продукты экзоцеллюлаз до моносахаров.
Оксидативные целлюлазы деполимеризуют целлюлозу радикальными реакциями. В случаи акцептора может выступать целлобиоза [21].
Целлюлитический комплекс грибов включает две большие группы:
1. Гидролазы, осуществляющие расщепление структурных полисахаридов клеточной стенки древесины, в том числе большая группа гликозилгидролаз, эндо- и экзополигалактуроназа и целый ряд других гликаназ и глюкозидаз, отщепляющих боковые остатки в основной цепи гемицеллюлаз и пектиназ. В это группу также включают некоторые эстеразы, помогающие гликозилгидролазам в расщеплении основных цепей структурных полисахаридов клеточной стенки (пектин-метилэстераза, ферулоилэстераза, ацетилксиланэстераза). Под гликозилгидролазой понимают ряд ферментов, такие как эндо- и экзоцеллюлаза, гемицеллюлаза, эндо- и экзоксиланазы, эндо- и экзоманназы, β-глюконаза.
2. Оксидоредуктазы, действующие на углеводные компоненты древесины. К ним относят глюкозооксидаза, галактозооксидаза, целлобиозодегидрогеназа [8].
Эндоглюканаза гидролизует β-1,4 связи между соседними остатками глюкозы в плотно упакованных областях целлюлозы, образуя разрывы в середине цепи.
Экзоглюканаза расщепляет разорванные целлюлозные цепи с нередуцирующих концов с образованием глюкозы, целлобиозы или целлотриозы.
Целлобиогидролаза отщепляет фрагмент из 10 остатков и более числа остатков глюкозы с нередуцирующих концов молекулы целлюлозы.
Целлобиоза или β-глюконаза катализирует превращение целлобиозы и целлотриозы в глюкозу [1].
Особый интерес среди внеклеточных ферментов представляют целлобиозодегидрогеназы. Это оксидаза, специфичная к окислению восстанавливающих сахаров 1,4-β-гликозидной связью (целлобиоза и целлоолигосахариды, маннобиоза и манноолигосахариды, лактоза), но не действующая на моносахариды и их олигомеры с α-глюкозидной связью, найдена (наряду с эндоцеллюлазами) во всех основных группах ксилотрофов, вызывающие мягкие, деструктивные и коррозионные гнили [4].
Фермент окисляет восстанавливающее звено подходящего ди- или олигосахарида до лактона, одновременно восстанавливая двух или одноэлектронный акцептор. Акцепторами могут быть хиноны и феноксирадикалы, образуемые лакказами или неспецифическими пероксидазами, либо ионы трехвалентного железа или двухвалентной меди. Принципиально возможно и каталитическое восстановление кислорода до перекиси водорода, хотя он является плохим субстратом целлобиозодегидрогеназы [4].
Ферментные системы мягких гнилей полностью разлагают упорядоченную целлюлозу в отсутствии кислорода, чем принципиально отличаются от ферментов возбудителей белой гнили, скорость действия которых на целлюлозу заметно снижалось при удалении растворенного кислорода. Обнаружение внеклеточной целлобиозодегидрогеназы у представителей термофильного микромицетаMyceliophthorathermophila предположили о возможности двух механизмов гидролазного и в присутствии ионов железа оксидазного, через свободнорадикальный процесс, запускаемый реактивом Фентона (Fe +2 +H2O2) в кислой среде [8].
Целлобиозодегидрогеназа имеет большое сродство к целлюлозе, может действовать на удаленном расстоянии, в глубине целлюлозных волокон, где образуется ее основной субстрат тАУ целлобиоза [8].
4. Практическое применение дереворазрушающих грибов и их
ферментов в биотехнологии
гриб разрушение древесина фермент биотехнология
Люди давно и широко используют грибы как продукт питания. Грибы богаты белками: в их сухом веществе 20-30% приходится на долю чистого белка. В них содержатся жиры, минеральные вещества, микроэлементы. К последним относят железо, кальций, цинк, йод, калий, фосфор. В сыроежках например, содержится 3,2% фосфора (от общего веса золы), в свинушке толстой
тАУ 9,3, в свинушке тонкой тАУ 3,9%. При этом шляпки содержат больше фосфора, чем ножки. В промышленных масштабах выращивают около 10 видов грибов, среди них такие как вешенка устричная и шиитаки. Грибы богаты витаминами и каратинойдамим, содержат немного жиров и уг
Вместе с этим смотрят:
Бiологiчне рiзноманiття людських рас
Бодрствование - нейрофизиологический процесс человека
Влияние биологически активных факторов окружающей среды на наследственные системы организма человека
Возрастная анатомия и физиология
Генетико-статистический анализ комбинационной способности сортов и форм яровой мягкой пшеницы по коэффициенту хозяйственной эффективности фотосинтеза