Хромосоми як матерiальна основа спадковостi. Змiни хромосом та механiзми iх реорганiзацii
Мiнiстерство освiти i науки Украiни Нацiональний технiчний унiверситет Украiни ВлКиiвський полiтехнiчний iнститутВ»
Факультет бiотехнологii i бiотехнiки
Реферат по курсу: Бiологiя клiтини
на тему: Хромосоми як матерiальна основа спадковостi. Змiни хромосом та механiзми iх реорганiзацii
Виконала:
студентка I курсу групи БТ-91
Терещук Ганна
Перевiрила:
Жолнер Л.Г.
Киiв 2009
Вступ
Успадкованi ознаки закладенi в матерiальних одиницях, генах, якi розташовуються в хромосомах клiтинного ядра. Хiмiчна природа генiв вiдома з 1944 г.: йдеться про дезоксирибонуклеiнову кислоту (ДНК). Фiзична структура була з'ясована в 1953 р. Подвiйна спiраль цiii макромолекули пояснюi механiзм спадковоi передачi ознак.
Придивляючись до навколишнього нас свiту, ми вiдзначаiмо велику рiзноманiтнiсть живих iстот тАУ вiд рослин до тварин. Рiзноманiтнiстю, що пiд цим здаiться, насправдi ховаiться дивна iднiсть живих клiтин тАУ елементiв, з яких зiбраний будь-який органiзм i взаiмодiiю яких визначаiться його гармонiйне iснування. З позицii вигляду схожiсть мiж окремими особинами велика, i все-таки не iснуi двох абсолютно iдентичних органiзмiв (не рахуючи однояйцевих близнят). Наприкiнцi XIX столiття в роботах Грегора Менделя були сформульованi основнi закони, що визначили спадкову передачу ознак з поколiння в поколiння. На початку ХХ столiття в дослiдах Т.Моргана було показано, що елементарнi успадкованi ознаки обумовленi матерiальними одиницями (генами), локалiзованими в хромосомах, де вони розташовуються послiдовно один за одним.
У 1944 р. роботи Еверi, Мак-Леода i Мак-Картi визначили хiмiчну природу генiв: вони складаються з дезоксирибонуклеiновоi кислоти (ДНК). Через 10 рокiв Дж. Уотсон i Ф. Крiк запропонували модель фiзичноi структури молекули ДНК. Довга молекула утворена подвiйною спiраллю, а комплiментарна взаiмодiя мiж двома нитками цiii спiралi дозволяi зрозумiти, яким чином генетична iнформацiя точно копiюiться (реплiцируiтся) i передаiться подальшим поколiнням.
Одночасно з цими вiдкриттями вченi намагалися проаналiзувати i ВлпродуктиВ» генiв, тобто тi молекули, якi синтезуються в клiтках пiд iх контролем. Роботи Ефрусси, Бiдла i Татума напередоднi другоi свiтовоi вiйни висунули iдею про те, що гени ВлпродукуютьВ» бiлки. Отже, ген зберiгаi iнформацiю для синтезу бiлка (ферменту), необхiдного для успiшного здiйснення в клiтцi певноi реакцii. Але довелося почекати до 60-х рокiв, перш нiж був розгаданий складний механiзм розшифровки iнформацii, увязненiй в ДНК, i ii переведення у форму бiлка. Врештi-решт, багато в чому завдяки працям Нiренберга (США), був вiдкритий закон вiдповiдностi мiж ДНК i бiлками тАУ генетичний код.
1. Структура ДНК
Ще в 1869 роцi швейцарський бiохiмiк Фрiдрiх Мiшер виявив в ядрi клiток з'iднання з кислотними властивостями i з ще бiльшою молекулярною масою, нiж бiлки. Альтман назвав iх нуклеiновими кислотами, вiд латинського слова ВлнуклеусВ» - ядро. Так само, як i бiлки, нуклеiновi кислоти i полiмерами. Мономерами iх служать нуклеотиди, у зв'язку з чим нуклеiновi кислоти можна ще назвати полiнуклеотидами.
Нуклеiновi кислоти були знайденi в клiтинах всiх органiзмiв, починаючи вiд простих i закiнчуючи вищими. Найдивнiше, що хiмiчний склад, структура i основнi властивостi цих речовин виявилися схожими у всiляких живих органiзмiв. Але якщо в побудовi бiлкiв беруть участь близько 20 видiв амiнокислот, то рiзних нуклеотидiв, що входять до складу нуклеiнових кислот, всього чотири.
У живих клiтинах мiститься два типи нуклеiнових кислот тАУ дезоксирибонуклеiнова (ДНК) i рiбонуклеiнова (РНК). Як ДНК, так i РНК несуть в собi нуклеотиди, що складаються з трьох компонентiв: азотистоi пiдстави, вуглеводу, залишку фосфорноi кислоти. Проте комбiнацiя цих компонентiв в ДНК i РНК декiлька рiзнi.
Фосфорна кислота в молекулах ДНК i РНК однакова. Вуглевод же i у двох варiантах: у нуклеотидiв ДНК тАУ дезоксирибоза, а в нуклеотидiв РНК тАУ рiбоза. РЖ рiбоза, i дезоксирибоза тАУ пятiчленниi, п'ятивуглецевi з'iднання тАУ пентози. В дезоксирибози, на вiдмiну вiд рiбози, лише на один атом кисню менше, що i визначаi ii назву, оскiльки дезоксирибоза в перекладi з латинського означаi позбавлена кисню рiбоза. Строга локалiзацiя дезоксирибози в ДНК, а рiбози в РНК, якраз i визначаi назву цих двох видiв нуклеiнових кислот.
Третiй компонент нуклеотидiв ДНК i РНК тАУ азотистi з'iднання, тобто речовини, що мiстять азот i що володiють лужними властивостями. У нуклеiновi кислоти входять двi групи азотистих основ. Однi з них вiдносяться до групи пiрiмiдiнiв, основу будови яких складаi шестичленне кiльце, а iншi до групи пуринiв, в яких до пiрiмiдiнового кiльця приiднано ще i пятiчленне кiльце.
До складу молекул ДНК i РНК входять два рiзнi пурини i два рiзнi пiримiдини. У ДНК i пурини тАУ аденiн, гуанiн i пiрiмiдiни тАУ цитозiн, тимiн. У молекулах РНК тi ж самi пурини, але з пiрiмiдiнов тАУ цитозiн i замiсть тимiну тАУ урацил. Залежно вiд вмiсту тiii або iншоi азотистоi пiдстави нуклеотиди називаються аденiловими, тiмiловимi, цитозiловимi, урациловимi, гуанiловимi.
Як же з'iднуються мiж собою нуклеотиди в довгi полiнуклеотiдниi ланцюги? Виявляiться, що таке з'iднання здiйснюiться шляхом встановлення зв'язку мiж залишком молекули фосфорноi кислоти одного нуклеотиду i вуглеводом iншого. Утворюiться сахаро-фосфорнiй скелет молекули полiнуклеотиду, до якого збоку один за iншим приiднуються азотистi основи.
Якщо врахувати, що в кожнiй нуклеiновiй кислотi по чотири види азотистих основ, то можна уявити собi безлiч способiв розташування iх в ланцюзi, подiбно до того, як можна в самiй рiзнiй послiдовностi нанизати на нитку намистинки чотирьох кольорiв тАУ червонi, бiлi, жовтi, зеленi. Послiдовнiсть розташування нуклеотидiв в ланцюгах молекул нуклеiнових кислот так само, як i амiнокислот в молекулах бiлкiв, строго специфiчна для клiток рiзних органiзмiв, тобто носить видовий характер.
ДНК i подвiйною спiраллю.
Полiнуклiiднi ланцюги досягають гiгантських розмiрiв. Цiлком зрозумiло, що у зв'язку з цим вони так само, як i бiлки, певним чином упакованi в клiтцi.
Модель структури молекули ДНК вперше створили бiохiмiки з Кембрiджського унiверситету в Англii Джеймс Уотсон i Френсис Крик. Було показано, що молекула ДНК складаiться з двох полiнуклеотiдних ланцюгiв, закручених одна довкола iншоi, з утворенням подвiйноi спiралi. Причому контакти iснують мiж обома полiнуклеотiднимi ланцюгами, точнiше, мiж пурином одного нуклеотиду i пiримiдином iншого. Отже зовнi молекулу ДНК можна представити як свого роду перекрученi мотузянi сходи.
Утворення зв'язкiв в молекулi ДНК тАУ процес строго закономiрний. Аденiловий нуклеотид може утворювати зв'язки лише з тiмiловим, а гуанiловий лише з цитозiловим. Ця закономiрнiсть отримала назву принципу комплiментарностi, тобто додатковостi. Насправдi, така строга послiдовнiсть у виборi пари наводить на думку, що в подвiйнiй молекулi ДНК аденiн як би доповнюi тимiн i навпаки, а гуанiн вiдповiдно тАУ цитозiн, як двi половинки розбитого дзеркала.
Принцип комплiментарностi дозволяi зрозумiти механiзм унiкальноi властивостi молекул ДНК тАУ iх здатнiсть самовiдтворюватися. ДНК тАУ це iдина речовина в живих клiтинах, що володii подiбною властивiстю. Процес самовiдтворення молекул ДНК вiдбуваiться при активнiй участi ферментiв. Особливi розплiтаючi бiлки послiдовно як би проходять уздовж системи водневих зв'язкiв, що сполучають азотистi основи обох полiнуклеотiдних ланцюгiв, i розривають iх. Одиночнi полiнуклеотiднi ланцюги ДНК, що утворилися в результатi, добудовуються згiдно з принципом комплiментарностi за допомогою ферменту за рахунок вiльних нуклеотидiв, що завжди знаходяться в цитоплазмi i ядрi. Навпроти гуанiлового нуклеотиду стаi вiльний цитозiловий нуклеотид, а навпроти цитозiлового, у свою чергу, гуанiловий i так далi. У ланцюзi, що знов утворився, виникають вуглеводно-фосфатнi i водневi зв'язки. Таким чином, в ходi самовiдтворення ДНК з однiii молекули синтезуються двi новi.
ДНК в клiтцi локалiзована в основному в ядрi, в його структурних компонентах тАУ хромосомах.
2. Хромосоми еукарiот
У 80-х роках минулого столiття в ядрах еукарiотичних клiтин були вiдкритi ниткоподiбнi структури (Ст Флеммiнг, Е. Страсбургер, Е. Ван Бенеден), названi В. Вальдейером (1888 р.) хромосомами (вiд греч. chroma тАУ колiр, забарвлення, soma тАУ тiло). Цим термiном була пiдкреслено сильна схожiсть хромосом в порiвняннi з iншими клiтинними органелами до основних фарбникiв. Протягом подальших 10 тАУ 15 рокiв бiльшiстю бiологiв було пiдтверджено, що саме хромосоми служать матерiальним носiiм спадковостi.
Хромосоми особливо чiтко видно пiд час дiлень клiтин, проте факт безперервностi iх iснування i в ядрах, що не дiляться, сумнiвiв не викликаi. Основна особливiсть функцiональних перетворень хромосом полягаi в циклi компактiзациi тАУ декомпактiзациi. У компактiзованном станi хромосомами i короткi товстi нитки, видимi в свiтловий мiкроскоп. В результатi декомпактiзациi хромосомна нитка стаi невидимою в свiтловий мiкроскоп, тому ядра багатьох живих клiтин виглядають оптично пустими.
Перетворення хромосом строго залежать вiд фаз клiтинного циклу, тому iх особливостi можуть розглядатися лише стосовно тiii або iншоi фази циклу. Промiжок часу мiж закiнченням одного клiтинного дiлення тАУ мiтоза i закiнченням подальшого називаiться мiтотичним циклом. Таким чином, мiтотичний цикл включаi мiтоз i промiжок мiж мiтозами тАУ iнтерфазу. РЖнтерфаза складаiться з трьох перiодiв: центрального тАУ фази синтезу ДНК (S), коли генетичний матерiал подвоюiться, а також передсинтетичного (G1) i синтетичного (G2) для поста, пiсля якого клiтина вступаi в мiтоз (М). Пiсля фази синтезу ДНК в G2-перiоде i в мiтозi, аж до анафази, в хромосомi виявляються двi нитки, званi сестринськими хроматидами.
Основний хiмiчний компонент хромосом тАУ молекули ДНК. Вмiст ii в ядрах соматичних клiток в два рази бiльше, нiж в ядрах зрiлих статевих клiток. Цi два типи клiтин вiдрiзняються один вiд одного i по числу хромосом. Число хромосом тАУ п в соматичних клiтках i кiлькiсть ДНК тАУ з (вiд англ. content тАУ вмiст) в них позначають як диплоiдне (2п хромосом, 2с ДНК), а в зрiлих статевих клiтинах як гаплоiдне (п хромосом, з ДНК).
2.1 Мiтоз
Мiтоз, або непряме дiлення, - основний спосiб розмноження еукарiотичних клiтин, що обумовлюi, зокрема, можливiсть збiльшення iх бiомаси, зростання i регенерацiю. Мiтоз складаiться з чотирьох фаз.

Перша тАУ профаза тАУ характеризуiться початком циклу компактизацii хромосом, який продовжуiться протягом всiii цiii фази. Внаслiдок цього хромосоми стають видимими пiд мiкроскопом, причому вже в середнiй профазi мiтоза вони представляються подвiйними структурами тАУ сестринськими хроматидами, закрученими одна довкола iншоi. До кiнця профази зникають ядерце i ядерна мембрана.
Друга тАУметафаза. Процес компактизацii хромосом продовжуiться i веде до ще бiльшого укорочення iх довжини. Хромосоми вишиковуються по екватору клiтки. Хроматиди сполученi мiж собою мiж собою в центромерi, званою також первинною перетяжкою. З'являються нитки мiтотичного веретена, якi приiднуються до ценромерiв. Кожна ценромера випробовуi напругу, оскiльки нитки веретена тягнуть ii до протилежних полюсiв.
Полюси клiтки формуються спецiальними органелами тАУ центросомами.
Третя тАУ анафаза тАУ починаiться з розриву ценромери, внаслiдок чого сестринськi хроматиди розходяться до рiзних полюсiв клiтки. З цiii митi кожна пара сестринських хроматид отримуi назву дочiрнiх хромосом.
Четверта тАУ телофаза. Хромосоми досягають полюсiв клiтки, з'являються ядерна мембрана, ядерце. Вiдбуваються декомпактiзация хромосом i вiдновлення структури iнтерфазного ядра. Закiнчуiться мiтоз дiленням цитоплазми i в типових випадках тАУ вiдновленням вихiдноi бiомаси дочiрнiх клiтин.
Бiологiчна роль мiтоза полягаi в забезпеченнi iдентичною генетичною iнформацiiю двох дочiрнiх клiток. Це досяжно лише завдяки циклу компактiзациi тАУ декомпактiзациi, який i дозволяi розподiлити спадковi молекули в мiнiмальному об'iмi мiтотичних хромосом. РЖнакше, враховуючи розмiри клiтки (десятки або сотнi кубiчних мiкрометрiв) i довжину декомпактiзованной хромосоми (сантиметри), кожне клiтинне дiлення супроводилося б хаотичним переплетенням хромосомного матерiалу.
У еволюцii еукарiотичних клiтин, мабуть, ця обставина i послужила причиною становлення настiльки складного генетичного процесу, як мiтоз.
2.2 Мейоз
Термiном ВлмейозВ» позначають два наступних один за одним дiлення, в результатi яких з диплоiдних клiток утворюються гаплоiдниi статевi клiтини, тАУ гамети. Якби заплiднення вiдбувалося диплоiдними гаметамi, то плодоiднiсть нащадкiв в кожному наступному поколiннi повинна була б зростати в геометричнiй прогресii. В той же час завдяки мейозу зрiлi гамети завжди гаплоiдни, що дозволяi зберiгати дiплоiдность соматичних клiток виду. Можливiсть iснування подiбного до мейозу дiлення при дозрiваннi гамет тварин i рослин була передбачена А. Вейсманом ще в 1887 р. Мейотичне дiлення не еквiвалентне мiтозу. Обом мейотичним дiленням передуi лише одна фаза синтезу ДНК.
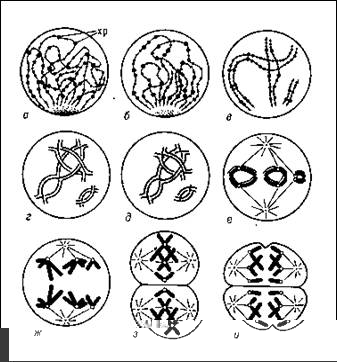
Тривалiсть ii, як i профази I дiлення мейозу, у багато разiв перевершуi вiдповiднi показники мiтотичного циклу будь-яких соматичних клiтин даного виду. Головнi подii мейозу розгортаються в профазi I дiлення. Вона складаiться з п'яти стадiй.
У першiй стадii тАУ лептотенi, слiдуючою безпосередньо за закiнченням предмейотичного синтезу ДНК, виявляються тонкi довгi хромосоми. Вони вiдрiзняються в профазi мiтоза двома особливостями: по-перше, в них не виявляiться подвiйнiсть, тобто не видно сестринських хроматид, по-друге, лептотеннi хромосоми мають виражену хромомерну будову. Хромомери тАУ вузлики. Дiлянки щiльноi компактiзациi ДНК, розмiри i розташування яких строго видоспецифiчний. Хромомери зустрiчаються як в мейотичних, так i в мiтотичних хромосомах, проте в останнiх без специфiчноi обробки вони не помiтнi.
Третя стадiя профази I дiлення тАУ пахитена тАУ у бiльшостi видiв найтривалiша. Пiд свiтловим мiкроскопом видно контАЩюгiровавшi хромосоми з бiльш менш чiтко вираженою хромомерною будовою. Приблизно в серединi пахитени мiж хроматидами гомологiчних хромосом з'являiться подовжня щiлина, яка чiтко показуi, що бiвалент тАУ це, по сутi, четверна хромосомна структура. У пахитене вiдбуваiться важлива генетична подiя тАУ кросинговер, або перехрещення хроматид гомологiчних хромосом. В результатi цього в кожному гомологу змiшуються батькiвський i материнський спадковий матерiал.
Результати кросинговера стають помiтними лише в четвертiй i п'ятiй стадiях профази I дiлення тАУ дiплотенi i дiакенезi. Дiплотена починаiться з моменту розбiжностi гомологiчних хромосом. В цей час в точках кросинговера видно перехрещенi хроматиди. Область перехрещення хроматид називають хiазмою. Число хiазм в цiлому вiдповiдаi кiлькостi актiв кросинговера в бiвалентi i пропорцiйно довжинi гомологiчних хромосом, його складових. Для дiплотени i дiакенезу характерне прогресуюче укорочення хромосом в результатi компактiзацii; тому хiазми поступово термiналiзуются, тобто наближаються до кiнцiв бiвалента i спадають з нього. Таким чином, у мiру наближення до метафази першого дiлення число хiазм зменшуiться.
У метафазi I дiлення мейозу район центромери кожноi хромосоми сполучений (на вiдмiну вiд метафази мiтоза) ниткою веретена лише з одним полюсом клiтки, причому центромери гомологiв, що розiйшлися, завжди пов'язанi з протилежними полюсами.
Анафазе I дiлення мейозу не передуi розщеплювання центромери, як при мiтозi, i тому до полюсiв вiдходять не хроматиди, а цiлi хромосоми, що складаються з двох хроматид. Проте, оскiльки гомологiчнi хромосоми розходяться до рiзних полюсiв, перше мейотичне дiлення приводить до редукцii числа хромосом. РЖншими словами, по числу хромосом продукти I дiлення мейозу стають гаплоiднимi. Проте у зв'язку з тим, що хромосоми в них зберiгають подвiйнiсть, тобто мiстять двi хроматиди, кiлькiсть ДНК зменшуiться лише до 2с.
Друге дiлення мейозу, наступне пiсля короткого промiжку тАУ iнтеркiнезу, приводить у вiдповiднiсть число хромосом i вмiст ДНК. Формально воно нагадуi мiтоз. На початку анафази вiдбуваiться роздiлення центромери, сестринськi хроматиди стають дочiрнiми хромосомами i розходяться до полюсiв. Таким чином, кожна з чотирьох клiтин, що утворилися унаслiдок двох мейотичних дiлень однiii клiтки, що пройшла предмейотичну s-фазу, мiститиме п хромосом i з ДНК.
Отже, головна вiдмiннiсть мейозу вiд мiтоза тАУ кон'югацiя гомологiчних хромосом з подальшою розбiжнiстю iх в рiзнi гамети. Точнiсть розбiжностi обумовлена точнiстю кон'югацii, а остання тАУ iдентичнiстю молекулярноi структури ДНК гомологiв.
2.3 Карiотип
Карiотипом називаiться хромосомний комплекс вигляду зi всiма його особливостями: числом хромосом, iх формою, наявнiстю видимих пiд свiтловим мiкроскопом деталей будови окремих хромосом. РЖнколи термiн ВлкарiотипВ» вживають по вiдношенню до хромосомного набору одиничноi клiтини або групи тканинних клiтин.
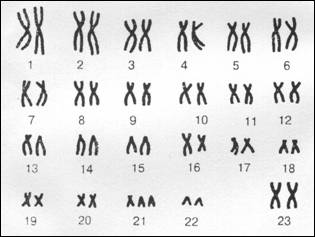
Число хромосом видоспецифiчне. Хоча закономiрностi, що характеризують карiотип, iнколи i вiдображають еволюцiю певних видiв, в цiлому по структурi карiотипу прямо судити про систематичне положення виду не можна.
У бiльшостi вищих тварин i рослин одна пара хромосом у особин однiii з родiв гетероморфна. Цi несхожi хромосоми називаються статевими. Зокрема, у ссавцiв i в дрозофiли клiтини чоловiчих органiзмiв мають Х- i y-хромосоми. В багатьох видiв y-хромосома вiдсутня. Всi останнi хромосоми називаються аутосомами.
Таким чином, завдяки дослiдженням цитологiв в кiнцi XIX тАУ початку ХХ ст була обгрунтована роль ядра в спадковостi, а спостереження за поведiнкою хромосом в мiтозе i мейозi привели до висновку, що саме з ними пов'язана передача спадкових ознак.
3. Секрети генетичного коду
У органiзмi кожноi людини тАУ своя спадкова конституцiя, характерна лише для нього. Саме з цим пов'язана тканинна несумiснiсть, що виявляiться, зокрема, при пересадцi органiв i тканин вiд одного органiзму iншому. ВлЧужаВ» шкiра, наприклад, зi своiми особливими молекулами вступаi в небажанi реакцii з органiзмом ВлгосподаряВ». Вона викликаi появу бiлкiв тАУ антитiл тАУ i в результатi не ВлприживаiтьсяВ». Аналогiчне явище спостерiгаiться i при пересадцi окремих органiв.
РЖнакше проходять цi процеси у однояйцевих близнят, якi розвиваються з двох клiтин, що утворилися з однiii заплiдненоi яйцеклiтини тАУ зiготи. Такi близнята завжди однополi i зовнi вражаюче схожi один на одного. У однояйцевих близнят пересадка тканин i органiв сповна можлива, жодного вiдторгнення iх не вiдбуваiться. РЖнакше i бути не може. Один i той же комплекс всiх спадкових чинникiв не провокуi появи антитiл в iх органiзмах.
Цi i багато iнших фактiв показали, що програмування синтезу бiлкiв тАУ головна властивiсть ДНК. Проте, перш нiж прийти до такого висновку, необхiдно було довести, що саме ДНК тАУ носiй генетичноi iнформацii. Перше пiдтвердження тому було отримано при вивченнi явищ трансформацii.
3.1 РЖсторiя доказу, що ДНК тАУ носiй генетичноi iнформацii
Явище це було вiдкрите в дослiдах з пневмококами, тобто з бактерiями, що викликають запалення легенiв. Вiдомо двi форми пневмококiв: А-форма з полiсахарiдной капсулою i б-форма без капсули. Обидвi цi ознаки спадковi.
Пневмококи а-форми при зараженнi ними мишей викликають запалення легенiв, вiд якого мишi гинуть. Б-форма для них нешкiдлива.
У 1928 роцi англiйський бактерiолог Ф.Грiффiтс заражав мишей сумiшшю, що складаiться з убитих нагрiванням пневмококiв а-форми i живих пневмококiв Б- форми. Учений передбачав, що мишi не захворiють. Але всупереч чеканням пiддослiднi тварини загинули. Ф. Грiффiтсу удалося видiлити з тканин загиблих мишей пневмококи. Всi вони виявилися капсулiрованнимi, тобто А- форми. Отже, убита форма якимсь чином передавала своi властивостi живим клiтинам б-форми. Але як?
Вiд рiшення цього питання залежало багато що, оскiльки, встановивши речовину, що передаi спадкову ознаку, тАУ утворення капсули, можна було отримати потрiбну вiдповiдь. Проте зробити це не виходило досить довго. Лише через 16 рокiв пiсля дослiдiв Ф. Грiффiтса, в 1944 роцi, американський учений А. Еверi iз спiвробiтниками, поставивши ряд чiтких експериментiв, зумiв з повним обгрунтуванням довести, що полiсахарид i бiлок не мають жодного вiдношення до передачi спадкових властивостей пневмокока а-форми.
В процесi цих експериментiв за допомогою спецiального ферменту розчинили полiсахаридну капсулу убитих пневмококiв а-форми i перевiрили, чи продовжують залишки клiтки форми А передавати спадкову iнформацiю клiтинам форми Б. Виявилось, що продовжують. Зрозумiло, що полiсахарид як джерело генетичноi iнформацii вiдпадаi.
Далi вченi за допомогою iнших ферментiв видалили iз залишкiв пневмококiв А бiлки i знову перевiрили iх дiю. Передача спадковоi iнформацii вiд А до Б продовжувалася. Отже, i бiлок нi при чому.
Таким чином, методом виключення було встановлено, що спадкову iнформацiю в клiтинi зберiгаi i передаi молекула ДНК. РЖ дiйсно, коли зруйнували ДНК, утворення капсульних форм А з беськапсульних Б припинилася.
Явище перетворення, тобто спадковоi змiни властивостей однiii форми бактерiй пiд впливом речовин iншоi форми, було назване трансформацiiю. Речовина ж, що викликаi трансформацiю, отримала назву трансформуючого агента. Ним, як було встановлено, служить ДНК.
3.2 Розшифровка генетичноi iнформацii
дезоксирибонуклеiновий хромосома генетичний спадковiсть
Полiмернi ланцюги бiлкiв складаються з мономiрних ланок тАУ амiнокислот i послiдовнiсть розташування iх в бiлковiй молекулi строго специфiчна. У зв'язку з цим очевидно, Що в ДНК повинна зберiгатися iнформацiя не лише про якiсний i кiлькiсний склад амiнокислот в молекулi даного бiлка, але i про послiдовнiсть iх розташування. Вiдповiдно якимсь чином мають бути закодованi в полiнуклеотидному ланцюзi ДНК кожна амiнокислота i бiлок в цiлому.
Знаючи, що амiнокислот всього 20, а нуклеотидiв тАУ 4, легко уявити собi, що 4 нуклеотида явно недостатньо для кодування 20 амiнокислот. Недостатньо також i коди з двох нуклеотидiв на кожну кислоту (4 = 16). Для кодування 20 амiнокислот необхiдно групи щонайменше з трьох нуклеотидiв (4 = 64). Подiбна група, що несе iнформацiю про одну амiнокислоту в молекулi бiлка, називаiться кодоном. Вся ж дiлянка ДНК, вiдповiдальна за синтез однiii молекули бiлка, в цiлому якраз i i ген. Значить, в генi стiльки кодонiв, скiльки амiнокислот входить до складу даного бiлка, що синтезуiться.
Синтез бiлкiв вiдбуваiться на рибосомах. ДНК же локалiзована в ядрi, в його хромосомах. Виникаi питання: яким чином генетична iнформацiя з ядра переноситься в цитоплазму на рибосому? Передбачити, що ДНК сама поступаi через пори ядерноi мембрани, не можна: Адже ДНК ядер володii величезною молекулярною масою i у зв'язку з цим просто не може проникнути через крихiтнi пори ядерноi мембрани. Тому мають бути якiсь дрiбнiшi молекули тАУ посередники, що передають генетичну iнформацiю вiд ДНК до бiлкiв. А.Н. Белозерський i А.Г. Спiрiн висунули мiркування, що цю роль грають молекули РНК.
Але одразу ж виникаi iнше питання: як копiюiться iнформацiя з ДНК на коротшi молекули РНК? Щоб вiдповiсти на нього, треба пригадати, що в будовi нуклеотиду ДНК i РНК багато загального. Зокрема, через схожiсть азотистих пiдстав iнформацiя з ДНК на РНК може переноситися за принципом комплiментарностi, згiдно з яким утворювати пари можуть не лише нуклеотиди в системi ДНК-ДНК, але i нуклеотиди в системi ДНК-РНК.
Оскiльки РНК так само, як i ДНК, мiстить пурiновi i пiрiмiдiновi основи, на дiлянках одного з ланцюгiв ДНК за допомогою ферменту РНК тАУ полiмерази будуються комплiментарнi короткi ланцюги РНК. Цей процес синтезу РНК на матрицi ДНК, що вiдбуваiться за допомогою ферментiв, носить назву транскрипцii. В результатi процесу транскрипцii закодована в ДНК послiдовнiсть нуклеотидiв, яка i i певною генетичною iнформацiiю, передаiться на РНК. Транскрипцiя вiдбуваiться на окремих дiлянках ДНК тАУ генах, кожен з яких мiстить набiр кодонiв, що програмують послiдовностi амiнокислот в данiй молекулi бiлка.
Рибонуклеiнова кислота, на якiй зроблена копiя ДНК, складаiться з одного ланцюга нуклеотидiв, в яких дезоксирибоза замiнена на рiбозу, а тимiн (Т) замiнений на урацил (У) .
Таким чином, в кожному кодонi ДНК транскрибiруiться в комплiментарний кодон РНК. В результатi виходить як би негатив РНК з позитиву тАУ ДНК. Ця РНК, що знiмаi iнформацiю з ДНК, називаiться iнформацiйнiй РНК (РЖ-РНК).
До теперiшнього часу ученим удалося розшифрувати кодони для всiх амiнокислот. Виявилось, що однiй амiнокислотi частенько вiдповiдаi декiлька кодонiв. Такий код називаiться виродженим. Поряд з цим виявилося, що деякi кодони не кодують жодну амiнокислоту. РЗх називають безглуздими. Безглуздi кодони мають дуже важливе значення, оскiльки визначають кордони початку i кiнця транскрипцii, тобто кордони генiв в данiй молекулi ДНК.
Якщо в прокарiот гени по своiму запису безперервнi, то в еукарiот це далеко не так. РЖнформацiя необхiдна для синтезу бiлка, виявляiться записаною з пропусками, переривчасто: гени складенi з кодуючих дiлянок (екзонiв), роздiлених некодуючими послiдовностями (iнтронами). При транскрипцii таких генiв iнтрони копiюються разом з екзонамi в загальну молекулу пре-мРНК. Остання пiддаiться в ядрi серii реакцiй, в ходi яких iнтрони вирiзуються, а екзони з'iднуються один з одним своiми краями. Молекула М-КОДУ-РНК, що вийшла, покидаi ядро i виявляiться вже у владi системи трансляцii, що дешифруi нуклеотидну послiдовнiсть. З'iднання амiнокислот з утворенням бiлка вiдбуваiться в цитоплазмi на особливих частках-рибосомах. Все це можна порiвняти з фабрикою (клiтина), в якiй креслення (гени) зберiгаються в бiблiотецi (ядро), а для випуску продукцii (бiлки) використовуються не самi креслення (ДНК), а iх фотокопiя (мРНК). Копiювальна машина (РНК-полiмераза) випускаi або по однiй сторiночцi фотокопii (ген), або вiдразу цiлу главу (оперон). Виготовленi копii видаються через спецiальнi вiконця (пори ядерноi мембрани). РЗх потiм використовують на монтажних лiнiях (рибосоми) з дешифратором (генетичний код) для здобуття iз заготовок (амiнокислот) остаточноi продукцii (бiлки).
Як же вiдбуваiться сам процес синтезу бiлка?
Перший його етап пов'язаний з функцiонуванням транспортноi РНК (т- РНК). Число рiзновидiв цих молекул РНК дорiвнюi числу основних амiнокислот, тобто iх 20 видiв. Кожнiй амiнокислотi вiдповiдаi певна Т-РНК i певний фермент.
У цитоплазмi клiтини завжди в достатнiй кiлькостi i рiзнi амiнокислоти. З них молекула Т-РНК вiдбираi вiдповiдну амiнокислоту. Кожна амiнокислота, перш нiж вступити в бiлковий ланцюг, за допомогою спецiального ферменту з'iднуiться з АТФ i запасаiться енергiiю.
ВлЗарядившисьВ» таким чином амiнокислота зв'язуiться з Т-РНК, яка переносить ii до рибосом. Характерною межею молекул Т-РНК i наявнiсть в iх структурах антикодонiв. Ця особливiсть забезпечуiться розташуванням вiдповiдних амiнокислот в тiй послiдовностi кодонiв, яка зашифрована в молекулi РЖ-РНК. Мiж рядом розташованими амiнокислотами виникають пептиднi зв'язки i синтезуiться молекула бiлка.
Таким чином, генетична iнформацiя, увтАЩязнена в ДНК, реалiзуiться рiзними видами РНК в молекулах вiдповiдних бiлкiв. Процес передачi програми, принесеноi з собою молекулами РЖ-РНК, отримав назву трансляцii.
4. Як же працюють гени?
Бiосинтез бiлкiв, що протiкаi пiд генетичним контролем, - це лише початок складних, багатоступiнчастих бiохiмiчних процесiв клiтки.
При вивченнi рослин, що вегетативно розмножуються, отриманi докази того, що окремi частини органiзму, такi як бульба, аркуш, цибулина, держак i так далi, дають початок нормальнiй рослинi. А це означаi, що всi клiтини даного органiзму несуть повну генетичну iнформацiю, так само як i вихiдна заплiднена яйцеклiтина, з якоi розвиваiться тварина. В той же час в будь-якому органiзмi мiстяться диференцiйованi клiтини з певною формою i функцiiю. Наприклад, у людини i клiтини нервовi, мтАЩязовi, статевi i так далi Але, не дивлячись на те, що кожна клiтина нашого тiла несе повну генетичну iнформацiю, тобто повний набiр генiв, отриманих вiд батькiв, функцiонують лише певнi гени, останнi знаходяться в неактивному станi. Яким же чином в клiтинi регулюiться дiяльнiсть тих або iнших генiв?
У всiх процесах життiдiяльностi клiтини роль бiологiчних каталiзаторiв грають ферменти. Без iх участi не протiкаi практично жодна хiмiчна реакцiя синтезу або розкладу речовин. У кожнiй клiтинi (з ii характерними функцiями) повиннi знаходитися регуляторнi механiзми, контролюючi не лише якiсний склад ферментiв, але i iх кiлькiсть. РЖнакше макромолекули бiлкiв, що безперервно синтезуються, накопичувалися б в клiтинi непотрiбним баластом, захаращуючи ii.
РЖ дiйсно, подiбний регуляторний механiзм був виявлений в клiтинах бактерiй в 1961 роцi французькими ученими Франсуа Жакобом i Жаком Моно.
Що ж це за механiзм?
Ф. Жакоб i Ж. Моно довели, що не всi гени бактерiй однаковi по своiму призначенню. Одна група тАУ структурнi гени, що видають iнформацiю про синтез певних полiпептидних ланцюгiв, iнша тАУ регуляторнi гени, що вiдають активнiстю структурних генiв шляхом iх ВлвключенняВ» i ВлвиключенняВ».
Регуляторнi гени представленi геном-оператором, безпосередньо зчепленим з групою структурних генiв, i геном-регулювальником, який може знаходитися в деякому вiддаленнi вiд них.
Ген-оператор з групою регульованих iм структурних генiв був названий опероном. Оперон служить одиницею транскрипцii, тобто з нього списуiться одна молекула РЖ-РНК. Ген регулювальник дii не шляхом безпосереднього контакту iз структурними генами, а за допомогою бiлка репрессора.
За наявностi молекул речовини, що синтезуiться, що досить накопичилися, белок-репрессор, з'iднуючись з цими молекулами, активiзуiться i зв'язуiться з геном-оператором. В результатi синтез даноi речовини припиняiться. Свою назву белок-репрессор отримав через те, що пригнiчуi дiяльнiсть гена-оператора, тобто ставить його в положення ВлвимкненоВ».
При малiй кiлькостi молекул, що синтезуються, бiлок-репрессор залишаiться неактивним. У таких умовах дiя оперону тАУ гена-оператора i структурних генiв тАУ не пригнiчуiться, i синтез продовжуватиметься безперешкодно.
5. Передача генетичноi iнформацii вiд батькiв до нащадкiв
Як вiдомо, особливостi, що характеризують нащадкiв, передаються iм вiд батькiв через статевi клiтини: чоловiчу тАУ сперматозоiд i жiночу тАУ яйцеклiтину. Злиття iх при заплiдненнi приводить до утворення iдиноi клiтини зiготи, з якоi розвиваiться зародок людини. Очевидно, що саме в цих двох статевих клiтинах i в зiготi, що утворилася при iх злиттi, зберiгаiться спадкова iнформацiя про фiзичнi, бiохiмiчнi i фiзiологiчнi властивостi, з якими з'являiться нова людина.
Матерiальною основою спадковостi служать нуклеiновi кислоти, а саме ДНК. Але яким же чином генетична iнформацiя передаiться вiд батькiв до потомства? Як вiдомо, новi клiтини з'являються в результатi дiлення вихiдних материнських.
Для бiльшостi клiтин характерне фiзiологiчно повноцiнне клiтинне дiлення, що складаiться з ряду фаз, пiд час яких ядро зазнаi закономiрних змiн, внаслiдок чого утворюються два ядра, абсолютно iдентичнi початковому. Цитоплазма при цьому дiлиться на двi половини. Таке складне дiлення отримала назва мiтоза, i характерний воно для клiтин тiла, тобто соматичних клiтин.
Проте, в органiзмах рослин, тварин i людини, окрiм соматичних, i i статевi клiтини. РЗх утворення вiдбуваiться в результатi особливого дiлення. Перетворення ж, яке викликаiться цим дiленням, отримало назву мейозу.
Пiд час i мiтозу i мейозу ядро втрачаi округлi контури i у ньому виразно вимальовувалися його структурнi компоненти, званi хромосомами. Хромосоми мають рiзнi форми: паличок, коротких стрижнiв, капiж i так далi.
Висновок
Вивчення генетики людини, незважаючи на всю складнiсть, важливе не лише з точки зору науки. Важко переоцiнити i прикладне значення дослiджень, що проводяться. Досягнення в цiй областi роблять помiтний вплив на iншi галузi наук про людину тАУ медицину, психiатрiю, психологiю, педагогiку.
Зокрема, велика роль генетики людини, що розвиваiться, у вирiшеннi проблем спадкових хвороб. Сучаснi данi свiдчать, що людиною успадковуються багато хвороб, таких, як не згортання кровi, колiрна слiпота, ряд психiчних захворювань. Крiм того, генетика людини покликана вирiшувати i iншi питання.
Значення розвитку генетики людини очевидно. Можна з повною упевненiстю сказати, що, наприклад, в молекулах ДНК клiток людини запрограмована генетична iнформацiя, контролююча кожну мить нашого життя. Це стосуiться здоров'я, нормального розвитку, тривалостi життя, спадкових хвороб, серцево-судинних захворювань, злоякiсних пухлин, схильностi до тих або iнших iнфе6кционним захворювань, старостi i навiть смертi.
Якщо видiлити з ядра однiii клiтини людини всi генетичнi молекули ДНК i розташувати iх в лiнiю одна за одною, то загальна довжина цiii лiнii складе сiм з половиною сантиметрiв. Така бiохiмiчна робоча поверхня хромосом. Це сконцентрована в молекулярному записi спадщина столiть минулоi еволюцii.
Правильно i образно сказав про це свого часу в романi ВлЛезо бритвиВ» письменник РЖван Ефремов: ВлСпадкова пам'ять людського органiзму тАУ результат життiвого досвiду незлiченних поколiнь, вiд риб'ячих наших предкiв до людини, вiд палеозойськоi ери до наших днiв. Ця iнстинктивна пам'ять клiтин i органiзму в цiлому i той автопiлот, який автоматично веде нас через всi прояви життя, борючись з хворобами, заставляючи дiяти складнi автоматичнi системи нервовою, хiмiчною, електричною i невiдомо якого ще регулювання. Чим бiльше ми взнаiмо бiологiю людини, тим бiльш складнi системи ми в нiй вiдкриваiмоВ».
Дослiдження останнiх рокiв довели, що будь-яка жива клiтина, у тому числi i клiтина людського органiзму, i цiлiсною системою, всi складовi елементи якоi виявляють тiсну взаiмодiю мiж собою i довкiллям, що робить на гени величезний вплив. Тому розрiзняють два поняття: генотип тАУ комплекс всiх спадкових фактiв тАУ генiв, що отримуються нащадками вiд батькiв, i фенотип тАУ сукупнiсть ознак, що виникають при взаiмодii генотипу i зовнiшнього середовища.
Отже, у формуваннi фенотипа важливi як генотип, так i зовнiшнi середовище, в якому вiдбуваiться розвиток особини. Без цiii взаiмодii неможливе було б життя, пов'язане з реалiзацiiю генетичноi iнформацii, увтАЩязненою в нуклеiнових кислотах.
Закономiрностi генетики в бiльшостi випадкiв носять унiверсальний характер. Вони однаково важливi для рослин, для тварин. Велике iх значення i для людини.
Список лiтератури
1. Генетика i спадковiсть. Збiрка статей. Г.34. Пер. з франц. М.: Свiт. 1987 р.
2. Загальна генетика. Алiхонян С. РЖ. РЖ iн. М.: Вища школа.1985 р.
3. Генетика вчора, сьогоднi i завтра. Дубiнiн Н.П. М.: Радянська Росiя. 1981р.
4. Бiологiчний словник енцеклопедiчеський.
5. Ботанiка: Морфологiя i анатомiя рослин. Васильiв А.Е. М.: Освiта. 1988 р.
Вместе с этим смотрят:
Бiологiчне рiзноманiття людських рас
Бодрствование - нейрофизиологический процесс человека
Влияние биологически активных факторов окружающей среды на наследственные системы организма человека
Возрастная анатомия и физиология