Загальна характеристика i особливостi життiдiяльностi бактерiй родини Enterobacteriaceae
Мiнiстерство освiти i науки Украiни
Днiпродзержинський державний технiчний унiверситет
Кафедра екологii та бiотехнологii
Дисциплiна Загальна мiкробiологiя
КУРСОВА РОБОТА
на тему
Загальна характеристика i особливостi життiдiяльностi бактерiй родини Enterobacteriaceae
Днiпродзержинськ
2009
ЗМРЖСТ
Вступ
1. Морфологiя бактерiй родини Enterobacteriaceae
2. Фiзiологiя бактерiй родини Enterobacteriaceae
3. Метаболiзм бактерiй родини Enterobacteriaceae
4. Генетика бактерiй родини Enterobacteriaceae
5. Екологiя бактерiй родини Enterobacteriaceae
6. Антигени бактерiй родини Enterobacteriaceae
7. Патогеннiсть i токсиноутворення бактерiй родини Enterobacteriaceae
8. Резистентнiсть бактерiй родини Enterobacteriaceae
9. Патогенез бактерiй родини Enterobacteriaceae
10. РЖмунiтет до бактерiй родини Enterobacteriaceae
11. Лабораторна дiагностика бактерiй родини Enterobacteriaceae
12. Профiлактика i лiкування захворювань викликаних бактерiями родини Enterobacteriaceae
13. Динамiка змiни резистентностi умовно-патогенних ентеробактерiй шлунково-кишкового тракту до антимiкробних препаратiв
Перелiк посилань
ВСТУП
Важливу роль в життiдiяльностi людини вiдiграi мiкрофлора товстого кишечника тАУ своiрiдний екстракопоральний орган. Вона i антагонiстом гнилiсноi мiкрофлори, так як продукуi молочну, оцтову кислоти, антибiотики i iнше. Вiдома ii роль в водно-сольовому обмiнi, регуляцii газового складу кишечнику, обмiнi бiлкiв, вуглеводiв, жирних кислот, холестерину i нуклеiнових кислот, а також продуктiв бiологiчно активних сполук тАУ антибiотикiв, вiтамiнiв, токсинiв та iн. Морфокiнетична роль мiкрофлори складаiться в ii участi в розвитку органiв i систем органiзму; вона приймаi участь також в фiзiологiчному запаленнi слизистоi оболонки i змiнi епiтелiю, перетравленнi i детоксикацii екзогенних субстратiв i метаболiтiв, що порiвнюiться з функцiiю печiнки. Нормальна мiкрофлора виконуi, крiм того, антимутагену роль, руйнуючи канцерогеннi речовини в кишечнику. В той же час деякi бактерii можуть продукувати потужнi мутагени. Пристiнкова мiкрофлора кишечника колонiзуi слизову оболонку у виглядi мiкроколонiй, утворюючи своiрiдну бiологiчну плiвку, яка складаiться з мiкробних тiл i екзополiсахаридного матриксу. Екзополiсахариди мiкроорганiзмiв, якi називаються глiкокалiксом, захищають мiкробнi клiтини вiд рiзних фiзико-хiмiчних i бiологiчних впливiв. Слизова оболонка кишечника також знаходиться пiд захистом бiологiчноi плiвки.
Великий вплив чинить мiкрофлора кишечника на формування i пiдтримку iмунiтету. В кишечнику мiститься близько 1,5 кг мiкроорганiзмiв, антигени яких стимулюють iмунну систему. Природнiм неспецифiчним стимулятором iмуногенезу i мурамiлдипептид, який утворюiться iз мiкрофлори пiд дiiю лiзоциму i iнших лiричних ферментiв, якi знаходяться у кишечнику.
Важливою функцiiю нормальноi мiкрофлори кишечнику i ii участь в колонiзацiйнiй резистентностi, пiд якою розумiють сукупнiсть захисних факторiв органiзму, i конкурентних, антагонiстичних i iнших особливостей анаеробiв кишечнику, якi надають стабiльнiсть мiкрофлорi i запобiгають колонiзацii слизових оболонок стороннiми мiкроорганiзмами [1].
До групи кишкових бактерiй вiдносяться дуже багато бактерii, зокрема родина Enterobacteriaceae. Це велика родина яка включаi такi роди як Salmonella, Escherichia, Klebsiella, Proteus, Shigella. Бiльшiсть бактерiй цiii родини складають значну частину нормальноi аеробноi флори кишечника i зазвичай не викликають захворювань, а навпаки, беруть участь у забезпеченнi його нормального функцiонування. Цi мiкроорганiзми стають патогенними лише у випадках проникнення iх у тканини органiзму, особливо в сечовидiльнi шляхи, легенi, очеревину чи мозковi оболонки, де вони здатнi викликати запальний процес. Вони можуть проникати в кров'яне русло (при використаннi венозних чи уретральних катетерiв); попадати в сечостатевi шляхи при проведеннi iнструментальних дослiджень (нозокомiаль-на передача); викликати iнфекцiйнi ураження сечовидiльних шляхiв пiсля введення контамiнованих лiкарських препаратiв, особливо пiсля внутрiшньовенного iхнього застосування. Можливий статевий шлях передачi.
Бактерii родини Enterobacteriaceae часто обумовлюють вториннi iнфекцii, у тому числi пневмонii, ендокардити (запалення внутрiшньоi оболонки серця), бактерiiмii, особливо в людей, що приймають наркотики, i в пацiiнтiв клiнiк.
Прояви iнфекцiй, викликаних бактерiями родини Enterobacteriaceae, залежать вiд локалiзацii патологiчного процесу. Вони не можуть бути диференцiйованi з iншими бактерiальними iнфекцiями на пiдставi одних лише симптомiв i ознак захворювання, тому дуже важливо визначити збудника iнфекцii, дати видову i типову характеристику, чутливiсть до антибактерiальних засобiв. Системне вивчення видових, типових i iнших особливостей мiкроорганiзмiв − збудникiв iнфекцiй − маi iстотне значення для з'ясування патогенезу iнфекцiй i визначення методiв iхньоi терапii.
Залежно вiд локалiзацii патологiчного процесу для дослiдження використовують видiлення статевих органiв, сечу, кров, спинномозкову рiдину та iн. У пофарбованих мазках виявляють грамнегативнi короткi палички, що можуть утворювати ланцюжки; вони дуже схожi один на одного i лише наявнiсть широких капсул маi дiагностичне значення для роду Klebsiella. Дослiджуваний матерiал одночасно наносять на кров'яний агар i середовища, що мiстять спецiальнi
барвники i вуглеводи, що дозволяi швидко диференцiювати ферганськi i неферганськi колонii. Бактерii, видiленi на цих середовищах, надалi iдентифiкують за допомогою бiохiмiчних i серологiчних реакцiй. Можлива швидка попередня iдентифiкацiя при можливостi мiкроорганiзмiв ферментувати лактозу.
Виражену протимiкробну дiю щодо бактерiй родини Enterobacteriaceae маi ампiцилiн, тетрациклiни, амiноглiкозиди, полiмiксини, сульфанiламiди. Однак чутливiсть до цих препаратiв в окремих штамiв рiзна, тому ii необхiдно визначати лабораторними методами. Нерiдко зустрiчаiться множинна стiйкiсть до лiкiв.
При лiкуваннi ефект може бути досягнутий при одночасному застосуваннi амiноглiкозидiв i фурагiну чи ампiцилiну; триметоприму iз сульфаметоксазолом i полiмiксином. Доцiльнi й iншi комбiнацii антибактерiальних засобiв, призначення препаратiв, що потенцiюють дiю антибiотикiв, що пiдвищують захиснi механiзми органiзму хворих, що сприяють доставцi антибактерiальних засобiв до вогнищ ураження й забезпечують iхню терапевтичну концентрацiю в тканинах i сироватцi кровi.
Профiлактика й ефективнiсть боротьби з цими iнфекцiями багато в чому залежать вiд дотримання чистоти рук, суворого дотримання правил асептики й антисептики (стерилiзацii iнструментарiю, дезiнфекцii устаткування), обережностi в призначеннi внутрiшньовенних препаратiв i особистоi гiгiiни сечостатевих шляхiв. Дуже важливо пам'ятати, що багато грамнегативних бактерiй i патогенними i викликають розвиток захворювання в ослабленому органiзмi. У лiкувальних установах цi бактерii нерiдко переносяться персоналом, а також через iнструменти чи препарати для парентерального введення [2].
1. МОРФОЛОГРЖЯ БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Мiкроорганiзми родини Enterobacteriaceae являють собою палички довжиною 1-5 мкм i товщиною 0,4-0,8 мкм. Вони добре фарбуються анiлiновими барвниками, грамнегативнi. Багато з них рухливi завдяки наявностi перетрихiально розташованих джгутикiв i можуть мати капсулу. Спор не утворюють. У багатьох штамiв, якi належать до рiзних родiв ентеробактерiй: ешерихiям, сальмонелам, шигелам, клебсiiлам, протеям, виявленi ворсинки (пiлi) загального типу, а у деяких штамiв тАУ статевi ворсинки (F-пiлi), з якими звтАЩязанi iх адгезивнi та iншi властивостi [3,4].
Ешерихii
Рiд E. coli представлений iдиним видом i багато чисельними бiоварами i сероварами, якi розрiзняються деякими бiохiмiчними, антигенними i патогенними властивостями.
Кишкову паличку (рис. 1.1) вперше видiлив iз фекалiй людини в 1885 роцi нiмецький педiатр i мiкробiолог Теодор Ешерих. В подальшому було встановлено, що подiбнi бактерii широко розповсюдженнi в природi. Вони зустрiчаються в кишечнику майже всiх видiв ссавцiв, птахiв, риб, а також в водi, ТСрунтi i iнших обтАЩiктах навколишнього середовища.
За своiми морфологiчними i тинкторiальними властивостями E. coli нагадуi iншi ентеробактерii. Це дрiбнi грамнегативнi палички довжиною 2-3 мкм, товщиною 0,5-0,7 мкм з закругленими кiнцями, в мазках розташовуються не упорядковано. Серед кишкових паличок зустрiчаються рухливi i не рухливi варiанти. Деякi штами мають виражену капсулу i утворюють слизовi колонii. S-форми E. coli на поживному агарi формують опалово-мутнуватi, слабко випуклi, вологi, з рiвними краями i блискучою поверхнею колонii, якi легко суспендуються в сольовому розчинi. Перехiднi S- i R-форми утворюють пласкi, з шершавою поверхнею i нерiвними краями колонii, якi погано суспендуються в сольовому розчинi. Кишкова паличка викликаi рiвномiрне помутнiння рiдини поживного середовища з утворенням невеликого осаду [3].
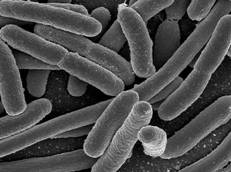
Рисунок 1.1 - Escherichia coli (за [5])
Сальмонели
До роду Salmonella (рис. 1.2) вiдносяться збудники черевного тифу, паратифу i сальмонельозiв. Назва роду походить вiд iменi американського дослiдника Данiеля Сальмона. Сальмонели пiдроздiленi на 4 пiдроду, якi включають 65 серогруп i близько 2000 сероварiв.
Сальмонели вiдповiдають в основному тим ознакам, якi присутнi всiм ентеробактерiям. При мiкроскопii мазкiв, забарвлених за грамом, черевнотифознi i паратифознi бактерii мають вигляд паличок червоного кольору iз заокругленими кiнцями розмiром 0,5-0,8×1-3 мкм, активно рухливi у висячiй чи роздавленiй краплi.
Рiст у МПБ супроводжуiться помутнiнням. На МПА виростають нiжнi, круглi, гладенькi, прозорi або напiвпрозорi колонii розмiром 2-4 мкм. Однак колонii тифозних мiкробiв, що мають Vi-антиген, каламутнi. У S. Schottmuelleri колонii грубiшi, через кiлька днiв по периферii колонii утворюiться слизовий валик.
На середовищах Ендо, Левiна, Плоскирiва колонii безбарвнi, прозорi, часом рожевуватi (Ендо) або злегка голубуватi (Левiна). На вiсмут-сульфатному агарi черевнотифознi мiкроби утворюють колонii чорного кольору, iнодi зi свiтлим обiдком. Паратифознi бактерii на цьому середовищi можуть утворювати коричневi або зеленуватi колонii. Пiсля зняття колонii залишаiться чорний слiд [3,6].
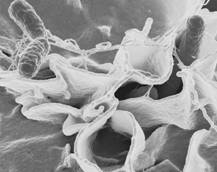
Рисунок 1.2 - Salmonella (за [7])
Шигели
Бактерii роду Shigella (рис. 1.3) i збудниками бактерiальноi дизентерii або шигельозу.
Першi припущення про роль бактерiй в етiологii дизентерii були зробленi А. Шентемесом i Ф. Вiдалем в 1888 роцi.
Дизентерiя тАУ полiетiологiчне захворювання. Його викликають рiзнi види бактерiй, якi називаються шигелами. В теперiшнiй час вони вiднесенi до роду Shigella, який пiдроздiляiться на чотири групи: A,B,C i D тАУ i бiльше двох десяткiв сероварiв.
За своiми морфологiчними властивостями шигел мало вiдрiзняються вiд ешерiхiй i сальмонел. Однак вони позбавленнi джгутикiв i тому i нерухомими бактерiями. Багато штамiв шигел мають ворсинки загального типу i статевi ворсинки. Рiзнi види шигел iдентичнi за своiми морфологiчними властивостями [3].
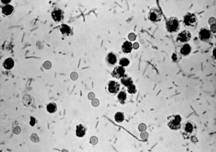
Рисунок 1.3 - Shigella (за [8])
Клебсiiли
У мазках забарвлених за Грамом, капсульнi форми клебсiiл мають вигляд грамнегативних, елiпсоподiбних, товстих паличок довжиною 5-8 мкм i шириною 3-5 мкм. Без капсульнi форми мають меншi розмiри (0,3-0,6×1-3 мкм), поодиноке, парне або ланцюжкове розташування [6].
Протей
Усi види роду Proteus (рис. 1.4) являють собою грамнегативнi прямi палички розмiрами 0,4-0,6×1,0-3,0 мкм. Зустрiчаються кокоподiбнi, нитковиднi форми. Спор i капсул не утворюють, мають перетрихiально розташованi джгутики [3].
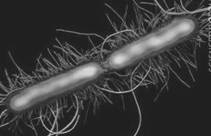
Рисунок 1.4 - P. vulgaris (за [9])
2. ФРЖЗРЖОЛОГРЖЯ БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Усi ентеробактерii i факультативними анаеробами, добре ростуть на середовищах з мтАЩясним екстрактом. Вони володiють вираженою ферментативною активнiстю, яка повтАЩязана з утворенням багато чисельних сахаридолiтичних, протеолiтичних i iнших ферментiв, як конституцiйних, так i iндуцибельних. Разом з тим вiдмiнностi якi спостерiгаються в перерахованих ознаках мають таксономiчне значення i використовуються для родинноi i видовоi диференцiацii i iдентифiкацii ентеробактерiй. Деякi з цих ознак слугують пiдставою для визначення окремих бiоварiв серед одного i того ж виду бактерiй.
До основних ознак, якi вказують на належнiсть ентеробактерiй до одного з птАЩяти триб вiдносяться:
1) характер продукту, який утворюiться при зброджуваннi глюкози: кислоти, бутандiол, iх сумiшi;
2) реакцiя з метиловим червоним;
3) утворення ацетилметилкарбiнолу i позитивна реакцiя Фогеса-Проскауера;
4) вiдновлення нiтратiв;
5) утворення уреази;
6) рiст в присутностi KCN i деякi iншi.
Головною бiохiмiчною ознакою, яка слугуi для визначення родовоi i видовоi належностi ентеробактерiй, i iх здатнiсть ферментувати рiзнi вуглеводи до утворення кислоти або газу, утворення iндолу, сiрководню, декарбоксилаз амiнокислот (лiзину, орнiтину та iн.), утилiзацiя цитрату, тартрату та iншi [3].
Ешерихii
Це факультативний анаероб, добре росте на звичайних поживних середовищах при слабо лужнiй реакцii середовища i оптимальнiй температурi 37В°С. Рiст i розмноження бактерiй можливi при доволi значних коливаннях рН середовища i температурного режиму.
Кишкова паличка маi найбiльш високу ферментативну активнiсть. Вона утилiзуi ацетат в якостi iдиного джерела вуглецю, вiдновлюi нiтрати до нiтритiв. Ферментуi глюкозу та iншi вуглеводи з утворенням пiрувату, який потiм перетворюiться в молочну, оцтову, мурашину кислоти видiляються CO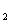 i H
i H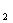 . Бiльшiсть штамiв ферментуi лактозу. Однак зустрiчаються штами, якi повiльно зброджують лактозу або зовсiм не володiють цiiю здатнiстю [3].
. Бiльшiсть штамiв ферментуi лактозу. Однак зустрiчаються штами, якi повiльно зброджують лактозу або зовсiм не володiють цiiю здатнiстю [3].
Сальмонели
Вони i ферментативно активними бактерiями, хоча деякi поступаються в цьому вiдношеннi ешерiхiям. Диференцiацiя багато чисельних представникiв даного роду проводиться на основi iх ферментативноi активностi i антигенноi структури наведено в таблицi 2.1 [3].
Таблиця 2.1 - Ферментативнi властивостi ешерихiй i тифозно-паратифозних бактерiй (за [6])
| Вид | Лактоза | Глюкоза | Мальтоза | Манiт | Сахароза | РЖндол | HS |
Escherichia coli Salmonella typhi Salmonella parathyphi A Salmonella schottmuelleri | кг - - - | кг к кг кг | кг к кг кг | кг к кг кг | кгВ± - - - | + - - - | В± + - + |
Примiтка: к-утворення кислоти; кг-утворення кислоти i газу; + - виявлення ознаки; − вiдсутнiсть ознаки; В± - можливе виявлення ознак.
Шигели
Збудники дизентерii хемоорганогетеротрофнi, невимогливi к поживним середовищам. На щiльних середовищах при видiленнi iз органiзму хворого утворюються, як правило, S-форми колонiй. Шигели виду sonnei утворюють два типи колонiй − S- (I фаза) i R- (II фаза). Бактерii I фази при пересiвах утворюють обидва типи колонiй. Оптимальна температура росту 37В°С, рН середовища 7,2.
Шигели менш ферментативно активнi, нiж iншi ентеробактерii: при зброджуваннi глюкози та iнших вуглеводiв утворюються кислi продукти без газоутворення.
Шигели не розщеплюють лактозу i сахарозу, за виключенням S. sonnei, якi повiльно (на 2-у добу) розщеплюють цi сахариди, наведено в таблицi 2.2 [3].
Таблиця 2.2 - Бiохiмiчнi властивостi шигел (за [6])
| Вид | Ферментацiя вуглеводiв | РЖндол | Каталаза |
| лактоза | глюкоза | мальтоза | манiт | дульцит | сахароза |
S. dysenteriae S. flexneri S. boydii S. sonnei | - - - + повiльно | + + + + | - + В± + | - + + + | - + + - | - - - + повiльно | - - + - | - - - + |
Клебсiiли
Вони невимогливi до поживного субстрату. Добре ростуть на основних поживних середовищах, утворюючи куполоподiбнi блискучi слизовi колонii. Викликають iнтенсивне помутнiння середовища при ростi у бульйонi.
Бiльшiсть штамiв ферментують глюкозу з утворенням кислоти i газу; використовують глюкозу i цитрат в якостi iдиного джерела вуглецю, а амiак - як джерело азоту.
Диференцiацiя рiзних видiв клебсiiл проводиться на основi iх рiзних здатностей ферментувати вуглеводи, утворювати уреазу i лiзиндекарбоксилазу, утилiзувати цитрат та iнших властивостей наведено в таблицi 2.3.
Клебсiiли i факультативними анаеробами. Оптимальна температура для iх росту 35-37В°С [3].
Таблиця 2.3 - Диференцiальнi ознаки клебсiiл (за [6])
| Вид | Ферментацiя | РЖндол | ВаРеакцiя Фогеса-Проскауера | ВаУтилiзацiя цитрату |
| Глюкоза | Лактоза | Сахароза | ВаСечовина |
K. pneumoniae K. oxytoca K. planticola K. terrigena K. ozaenae K.rhinoskleromatis | + + + + В± В± | + + + + В± − | + + + + + В± | + + + - - - | - + В± - - - | + + + + - - | + + + В± В± - |
Протеi
Вони не вимогливi до поживного субстрату, добре ростуть на основних поживних середовища. На середовищах Ендо i Плоскирiва виростають прозорi блискучi безбарвнi колонii. На вiсмут-сульфатному агарi через 48 годин утворюються сiрувато-коричневi колонii, пiд якими формуiться чорно-коричнева зона. Н-форма (джгутикова) на простому агарi даi характерний повзучий рiст або тАЮфеномен роiннятАЭ. Культура маi неприiмний гнильний запах. При посiвi у стовпчик напiврiдкого агару визначають рухливiсть. Повзучий характер росту протею використовують для видiлення чистих культур посiвом у конденсацiйну воду скошеного агару за методом Шукевича. Протеi i факультативними анаеробами. Ростуть при температурi 20-37В°С.
Ферментують багато вуглеводiв з утворенням кислих продуктiв, глюкозу розщеплюють з утворенням кислот i невеликоi кiлькостi газу. Рiзнi види вiдрiзняються один вiд одного за ферментацiiю вуглеводiв, утворенню iндолу, уреази, сiрководню, орнiтиндекарбоксилази та iншим ознакам, наведено в таблицi 2.4 [3,6].
Таблиця 2.4 - Диференцiальнi ознаки бактерiй роду Proteus (за [6])
| Вид | Ферментацiя | Утворення | Орнiтин-декарбокси- лаза | ВаРеакцiя Фогеса-Проскауера |
| мальтози | сахарози | ксилози | iндолу | HS |
P. vulgaris P. mirabilis P. myxofaciens P. penneri | + - + + | + - + + | + + - + | + - - - | + + - В± | - + - - | - В± + - |
| | | | | | | | | | |
3. МЕТАБОЛРЖЗМ БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Бiльшiсть ентеробактерiй можуть використовувати в якостi субстратiв для дихального метаболiзму велику кiлькiсть простих органiчних сполук тАУ органiчнi кислоти, амiнокислоти, вуглеводи. В аеробних умовах, всi цi бактерii добре ростуть на звичайних складних бактерiологiчних середовищах, азотистi компоненти яких (амiнокислоти i пептиди) забезпечують iх окислюючими субстратами. В анаеробних умовах, однак, рiст стаi строго залежним вiд наявностi зброджуi мого вуглеводу. Деякi моносахариди, дисахариди i полiспирти зброджуються всiма представниками даноi групи. Використання полiсахаридiв менш розповсюджено.
Хоча ентеробактерii вирощують на складних середовищах, мiнiмальнi харчовi потреби цих органiзмiв, зазвичай простi. Представники багатьох родiв взагалi не потребують факторiв росту (наприклад Escherichia coli, багато видiв Salmonella), а у ауксотрофних органiзмiв цi потреби зазвичай простi. Особливо розповсюджена потреба в нiкотиновiй кислотi, що характерна для багатьох видiв роду Proteus. У Salmonella typhi i специфiчна потреба в триптофанi.
Дослiдження механiзму регуляцii бiосинтезу амiнокислот у багатьох ентеробактерiй виявило характернi особливостi цього процесу, якi вiдрiзняють даннi органiзми вiд всiх iнших бактерiй. Наприклад, перша стадiя в бiосинтезi амiнокислот родини аспартату, перетворення аспарагiновоi кислоти в аспартилфосфат, у ентеробактерiй завжди здiйснюiться трьома iзофункцiональними аспартаткiназами: аспартаткiназою I, яка i iнгiбуiться, i репресуiться треонiном, аспартаткiназою II, яка репресуiться, але не iнгiбуiться метiонiном, i аспартаткiназою III, яка i iнгiбуiться, i репресуiться лiзином. Цей тип регуляцii аспарагiнового шляху не був виявлений нi в однiii бактерii не кишковоi групи.
ГЦ-склад ДНК ентеробактерiй варiюi в широких межах тАУ вiд 37 до 63%. За виключенням роду Proteus, представники якого суттiво вiдрiзняються за нуклеотидним складом. ГЦ-склад ДНК спорiднених органiзмiв трьох видiв - Salmonella, Escherichia i Shigella тАУ розрiзняються зовсiм мало. Дiапазон змiнення
ГЦ-складу для тАЮкласичнихтАЭ бактерiй родини Enterobacteriaceae (вiд 37 до 59%) дуже близький до дiапазону для представникiв родини з полярним джгутикуванням (вiд 39 до 63%) [10].
Бродiння
Зброджування сахаридiв у ентеробактерiй вiдбуваiться за шляхом Ембдена-Мейiргофа. Продукти бродiння у рiзних представникiв групи розрiзняються i якiсно, i кiлькiсно. Однак у цих процесiв i одна характерна бiохiмiчна властивiсть, яка рiдко зустрiчаiться при iнших типах бактерiального бродiння. Мова йде про особливий спосiб розщеплення однiii з промiжних сполук тАУ пiровиноградноi кислоти (ПВК), що веде до утворення мурашиноi кислоти:
СН3СОСООН + КоАSН→СН3СОSКоА+ НСООН.
Таким чином, мурашина кислота часто i основним кiнцевим продуктом бродiння. Вона накопичуiться, хоча не завжди, оскiльки деяки з цих бактерiй володiють формнатгiдрогенлiазою, яка розщеплюi мурашину кислоту до СО2 i Н2:
НСООН → СО2+Н2.
У таких органiзмiв замiсть мурашиноi кислоти як кiнцевого продукту бродiння утворюються еквiмолярнi кiлькостi Н2 i СО2.
Найбiльш розповсюдженим типом перетворення сахаридiв при бродiннi у ентеробактерiй i так зване бродiння змiшаного типу, яке приводить до утворення молочноi, оцтовоi, бурштиновоi i мурашиноi кислот (або СО2 i Н2), а також етилового спирту. Такий тип бродiння властив представникам родiв Salmonella, Escherichia, Shigella i Proteus. Вiдношення кiлькостi рiзних кiнцевих продуктiв рiзниться як вiд штаму до штаму, так i для одного штаму при ростi в рiзних умовах, наприклад при рiзних значеннях рН. Така рiзниця вiдображаi ту обставину, що кiнцевi продукти утворюються з пiровиноградноi кислоти трьома незалежними шляхами (рис. 3.1). Утворення газу в результатi зброджування сахаридiв i дуже важливою ознакою для ентеробактерiй тАУ газоутворювальнi бактерii роду Escherichia вiдрiзняються за цим критерiiм вiд патогенних видiв групи Shigella i Salmonella typhi, якi зброджують сахариди без видiлення газу. В випадку простого бродiння змiшаного типу газ може утворюватися тiльки за рахунок розщеплення мурашиноi кислоти; вiдповiдно, утворення газу вiдображаi наявнiсть формiатгiдрогенлiази. Цей ферментний комплекс, звiсно, не суттiвий для бродiння i може загубитися в результатi мутацii без змiни здатностi бактерii до бродiння. РЖ дiйсно, дослiд показуi, що в природi iснують тАЮнеаерогеннiтАЭ, тобто не утворюючi газоподiбних продуктiв штами такого типового продуценту iх, як Escherichia coli. Тому, хоча видiлення газу являi собою корисну ознаку при класифiкацii ентеробактерiй, даний критерiй не i безпомилковим.
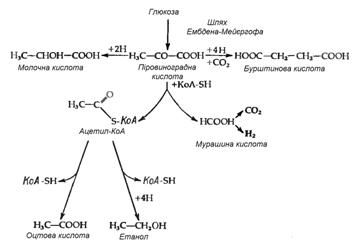
Рисунок 3.1 - Шляхи утворення iз ПВК типових кiнцевих продуктiв кислотного бродiння змiшаного типу (за [10])
РЖншою ознакою, яка маi велике значення для iдентифiкацii ентеробактерiй, i здатнiсть зброджувати дисахарид лактозу, що обумовлено наявнiстю у цих органiзмiв β-галактозiдази. Ефективнiсть зброджування лактози залежить ще i вiд наявностi специфiчноi пермеази галактозидiв, яка сприяi проникненю лактози в клiтину. Штами якi мають β-галактозiдазу, але позбавленнi пермеази, не можуть поглинати лактозу зi швидкiстю, яка достатня для iнтенсивного бродiння, i зазвичай класифiкуються як не здатнi до зародження цього сахариду. Зброджування лактози властиво Escherichia, але вiдсутнi у Salmonella, Shigella i Proteus. Деякi штами Shigella утворюють β-галактозiдазу, але не можуть зброджувати лактозу iз-за вiдсутностi пермеази [10].
4. ГЕНЕТИКА БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Вiдкриття явища переносу генiв у ентеробактерiй при контАЩюгацii i трансдукцii дозволило доволi детально дослiдити генетичнi родиннi вiдносини мiж деякими ii представниками. Утворення хромосомних гiбридiв мiж E. coli i бактерiями родiв Salmonella i Shigella каже про дуже високу ступiнь генетичноi гомологii цих бактерiй, що пiдтверджуiться i порiвнянням хромосомних карт двох найбiльш вивчених в цьому вiдношеннi видiв − Salmonella typhimurium i Escherichia coli. Як видно на рис. 4.1 i 4.2, де представленнi в спрощеному виглядi хромосомнi карти цих двох органiзмiв, багато маркерiв розташовуються на обох хромосомах в однакових локусах.
Про близьку генетичну спорiдненiсть груп Escherichia, Salmonella i Shigella свiдчить також висока ступень ДНК-ДНК-гiбридизацii in vitro. Утворення ж хромосомних гiбридiв мiж бактерiями цiii пiдгрупи i представниками iнших родiв ентеробактерiй (Proteus, Enterobacter) вiдбуваiться дуже рiдко. Крiм того, випробування по гiбридизацii ДНК-ДНК вказують, що ступень генетичноi гомологii мiж представниками пiдгрупи Escherichia− Salmonella− Shigella та iншими групами ентеробактерiй доволi низька. Ентеробактерii, що належать до рiзних родiв, можуть отримувати плазмiни вiд донорних штамiв E. coli при контАЩюгацii i потiм зберiгати iх як позахромосомнi елементи (ефективнiсть контАЩюгацii варiюi в широких межах; таблиця 4.1). Таким чином серед бактерiй кишковоi групи можуть розповсюджуватися F-фактори, якi мiстять додатковi гени (наприклад F-lac), i R-фактори, якi визначають стiйкiсть до рiзноманiтних лiкарських засобiв [10].
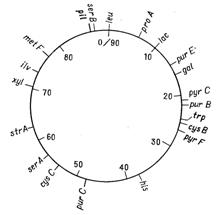
Рисунок 4.1 - Генетична карта E. coli К12 в спрощеному виглядi (за [10])
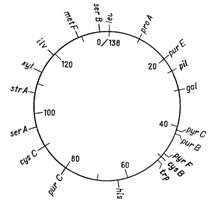
Рисунок 4.2 - Генетична карта S. typhimurium в спрощеному виглядi (за [10])
5. ЕКОЛОГРЖЯ БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Середовищем проживання для бiльшостi ентеробактерiй i кишковий тракт хребетних тварин i людини.
В органiзмi людини багато ентеробактерiй мiстяться в складi мiкробних бiоценозiв тонкого i товстого кишечника. Патогеннi види зустрiчаються тiльки у хворих i бактерiоносiiв.
З випорожненнями людей i тварин ентеробактерii потрапляють в навколишнi середовище. В нiй вони можуть зберiгатися на протязi рiзних строкiв в залежностi вiд виду i умов. Деякi з них (E. coli) використовуються в санiтарнiй мiкробiологii в якостi показникiв фекального забруднення навколишнього середовища [3].
Кишкова паличка наприклад може зберiгатися в водi i ТСрунтi декiлька мiсяцiв [11].
Широке розповсюдження сальмонел в природi i рiзноманiття шляхiв, якими вони проникають в органiзм людини, пояснюiться iх генетичною пластичнiстю. Сальмонели легко знаходять екологiчнi нiшi i адаптуються до найрiзноманiтнiших умов [12].
Клебсiiли тАУ широко розповсюдженi бактерii. Вони порiвняно стiйкi до факторiв навколишнього середовища завдяки наявностi капсули i можуть на протязi тривалого часу зберiгатися в ТСрунтi, водi, примiщеннях. Разом з тим вони зустрiчаються в складi мiкробних бiоценозiв в органiзмi людини i тварин [3].
Протеi P. vulgaris i P. mirabilis i мешканцями кишечника багатьох тварин, виявляються також в стiчних водах i ТСрунтi. Всi види протеiв можуть видiлятися з хворих людей [3].
6. АНТИГЕНИ БАКТЕРРЖЙ РОДИНИ ENTEROBACTERIACEAE
Антигенна будова служить одним iз суттiвих критерiiв, на яких основана класифiкацiя, а також iдентифiкацiя ентеробактерii. Розрiзняють три основних типи антигенiв: 1) О-соматичний антиген; 2) Н-джгутиковий антиген; 3)К-антигени. О-антиген i складовою частиною лiпополiсахариду (ЛПС) зовнiшнього шару клiтинноi стiнки. Специфiчнiсть О-антигену визначаiться детермiнантними сахаридами (гексозами i амiносахаридами), ковалентно звтАЩязаними з базисною частиною ЛПС. Н-антиген локалiзуiться в джгутиках клiтини. Вiн складаiться з бiлка флагелiну. Капсульнi антигени К-антигени, так як i О-антигени, мiстяться в ЛПС клiтинноi стiнки, але в бiльш поверхневому шарi. Вони маскують О-антигени.
В звтАЩязку з цим для визначення О-антигену у бактерiй, якi володiють К-антигеном, необхiдно зруйнувати останнiй шляхом киптАЩятiння або автоклавування культури. К-антигени за хiмiчними властивостями вiдносяться до кислих полiсахаридiв. До К-антигенiв вiдносяться К-антигени E. coli, М-антигени Salmonella schottmuelleri, Vi-антиген збудника черевного тифу, який виявляiться також у S. parathyphi C та деяких штамiв E. coli. Всi цi антигени характеризуються iмуннохiмiчною специфiчнiстю, що дозволяi диференцiювати роди i види, а також видiляти серед них серогрупи i серологiчнi варiанти (серовари). Антигенну будову е
Вместе с этим смотрят:
Бодрствование - нейрофизиологический процесс человека
Генетична iнформацiя
Естествознание в системе наук
Изменчивость организмов
Классификация и строение водорослей