Зрительная система
СОДЕРЖАНИЕ
1 ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЗРИТЕЛЬНОЙ СЕНСОРНОЙ СИСТЕМЫ
1.1 Основные показатели зрения
1.2 Психофизические характеристики света
1.3 Периферический отдел зрительной системы
2 СОМАТОВИiЕРАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
2.1 Психофизика кожной механорецепции
2.2 Кожные механорецепторы
2.3 Психофизика терморецепции
2.4 Терморецепторы
2.5 Висцеральная чувствительность
2.6 Проприоцепция
2.7 Функциональный и анатомический обзор центральной соматосенсорной системы
2.8 Передача соматовисцеральной информации в спинном мозгу
2.9 Соматосенсорные функции ствола мозга
2.10 Таламус
2.11 Соматосенсорные проекционные области в коре
2.12 Контроль афферентного входа в соматосенсорной системе
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1 ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЗРИТЕЛЬНОЙ СЕНСОРНОЙ СИСТЕМЫ
Зрительная система (зрительный анализатор) представляет собой совокупность защитных, оптических, рецепторних и нервных структур, воспринимающих и анализирующих световые раздражители. В физическом смысле свет тАФ это электромагнитное излучение с различными длинами волн тАФ от коротких (красная область спектра) до длинных (синяя область спектра).
Способность видеть объекты связана с отражением света от их поверхности. Цвет зависит от того, какую часть спектра поглощает или отражает предмет. Главные характеристики светового стимула тАФ его частота и интенсивность. Частота (величина, обратная длине волны) определяет окраску света, интенсивность тАФ яркость. Диапазон интенсивностей, воспринимаемых глазом человека тАФ огромен тАФ порядка 1016. Через зрительную систему человек получает более 80% информации о внешнем мире.
1.1 Основные показатели зрения
Зрение характеризуют следующие показатели:
1) диапазон воспринимаемых частот или длин волн света;
2) диапазон интенсивностей световых волн от порога восприятия до болевого порога;
3) пространственная разрешающая способность тАФ острота зрения;
4) временная разрешающая способность тАФ время суммации и критическая частота мельканий;
5) порог чувствительности и адаптация;
6) способность к восприятию цветов;
7) стереоскопия тАФ восприятие глубины.
1.2 Психофизические характеристики света
Психофизические эквиваленты частоты и интенсивности света представлены в таблицах 1.1 и 1.2.
Таблица 1.1. Психофизические эквиваленты частоты света

Таблица 1.2. Психофизические эквиваленты интенсивности света
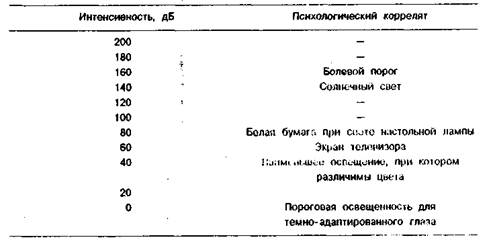
Для характеристики восприятия света важны три качества: тон, насыщенность и яркость. Тон соответствует цвету и меняется с изменением длины волны света. Насыщенность означает количество монохроматического света, добавление которого к белому свету обеспечивает получение ощущения, соответствующего длине волны добавленного монохроматического света, содержащего только одну частоту (или длину волны). Яркость света связана с его интенсивностью. Диапазон интенсивностей света от порога восприятия до величин, вызывающих болевые ощущения, огромен тАФ 160 дБ. Воспринимаемая человеком яркость объекта зависит не только от интенсивности, но и от окружающего его фона. Если фигура (зрительный стимул) и фон освещены одинаково, то есть между ними нет контраста, яркость фигур возрастает с увеличением физической интенсивности освещения. Если контраст между фигурой и фоном увеличивается, яркость воспринимаемой фигуры уменьшается с увеличением освещенности.
Пространственная разрешающая способность тАФ острота зрения тАФ минимальное различимое глазом угловое расстояние между двумя объектами (точками). Острота определяется с помощью специальных таблиц из букв и колец и измеряется величиной I/a, где а тАФ угол, соответствующий минимальному расстоянию между двумя соседними точками разрыва в кольце. Острота зрения зависит от общей освещенности окружающих предметов. При дневном свете она максимальна, в сумерках и в темноте острота зрения падает.
Временные характеристики зрения описываются двумя основными показателями тАФ временем суммации и критической частотой мельканий.
Зрительная система обладает определенной инерционностью: после включения стимула необходимо время для появления зрительной реакции (оно включает время, требующееся для развития химических процессов в рецепторах). Исчезает зрительное впечатление не сразу, а лишь через некоторое время после прекращения действия на глаз света или изображения, поскольку для восстановления зрительного пигмента сетчатке глаза также требуется время. Существует эквивалентность между интенсивностью и длительностью действия света на глаз. Чем короче зрительный стимул, тем большую интенсивность он должен иметь, чтобы вызывать зрительное ощущение. Таким образом, для возникновения зрительного ощущения имеет значение суммарное количество световой энергии. Эта связь между длительностью и интенсивностью сохраняется лишь при коротких длительностях стимулов тАФ до 20 мс. Для более длительных сигналов (от 20 мс до 250 мс) полная компенсация пороговой интенсивности (яркости) за счет длительности уже не наблюдается. Всякая зависимость между способностью к обнаружению света и его длительностью исчезает после того, как продолжительность стимула достигает 250 мс, а при больших длительностях решающей становится интенсивность. Зависимость пороговой интенсивности света от длительности его воздействия называется временной суммацией. Этот показатель используется для оценки функции зрительной системы.
Зрительная система сохраняет следы светового раздражения в течение 150-250 мс после его включения. Это свидетельствует о том, что глаз воспринимает прерывистый свет, как непрерывный, при определенных интервалах между вспышками. Частота вспышек, при которой ряд последовательных вспышек воспринимается как непрерывный свет, называется критической частотой мельканий. Этот показатель неразрывно связан с временной суммацией: процесс суммации обеспечивает плавное слияние последовательных изображений в непрерывный поток зрительных впечатлений. Чем выше интенсивность световых вспышек, тем выше критическая частота мельканий. Критическая частота мельканий пи средней интенсивности света составляет 16-20 в 1 с.
Порог световой чувствительности тАФ это наименьшая интенсивность света, которую человек способен увидеть. Она составляет 10 -10 тАФ 10 -11 эрг/с. В реальных условиях на величину порога существенно влияет процесс адаптации тАФ изменения чувствительности зрительной системы в зависимости от исходной освещенности. При низкой интенсивности света в окружающей среде развивается темповая адаптация зрительной системы. По мере развития темновой адаптации чувствительность зрения возрастает. Длительность полной темновой адаптации составляет 30 мин. При увеличении освещенности окружающей среды происходит световая адаптация, которая завершается за 15-60 с. Различия темновой и световой адаптации связаны со скоростью химических процессов распада и синтеза пигментов сетчатки.
Восприятие света зависит от длины волны света, попадающего в глаз. Однако, такое утверждение справедливо лишь для монохроматических лучей, то есть лучей с одной длиной волны. Белый свет содержит все длины световых волн. Существует три основных цвета: красный тАФ 700 нм, зеленый тАФ 546 нм и синий тАФ 435 нм. В результате смешивания основных цветов можно получить любой цвет. Объясняют цветовое зрение на основе предположения о существовании в сетчатке глаза фоторецепторов трех различных типов, чувствительных к различным длинам волн света, соответствующих основным частотам спектра (синий, зеленый, красный).
Нарушение восприятия цвета называется цветовой слепотой, или дальтонизмом, по имени Дальтона, который впервые описал этот дефект зрения на основе собственного опыта. Дальтонизмом страдают, в основном, мужчины (около 10%) в связи с отсутствием определенного гена в Х-хромосоме. Известны три типа нарушений светового зрения: протанопия тАФ отсутствие чувствительности к красному цвету, дейтеранопия тАФ отсутствие чувствительности к зеленому цвету и тританопия тАФ отсутствие чувствительности к синему цвету. Полная цветовая слепота тАФ монохроматия тАФ встречается исключительно редко.
Бинокулярное зрение тАФ участие обоих глаз в формировании зрительного образа тАФ создается за счет объединения двух монокулярных изображений объектов, усиливая впечатление пространственной глубины. Поскольку глаза расположены в разных "точках" головы справа и слева, то в изображениях, фиксируемых разными глазами, имеются небольшие геометрические различия (диспарантность), которые тем больше, чем ближе находится рассматриваемый объект. Диспарантность двух изображений лежит в основе стереоскопии, то есть восприятия глубины. Когда голова человека находится в нормальном положении, возникают отклонения от точно соответствующих проекций изображений в правом и левом глазах, так называемая диспарантность рецептивных полей. Она уменьшается с увеличением расстояния между глазами и объектом. Поэтому на больших расстояниях между стимулом и глазом глубина изображения не воспринимается.
1.3 Периферический отдел зрительной системы
Снаружи глаз виден как сферическое образование, прикрытое верхним и нижним веком и состоящее из склеры, коньюктивы, роговицы, радужной оболочки. Склера представляет собой соединительную ткань белого цвета, окружающую глазное яблоко. Коньюктива тАФ прозрачная ткань, снабженная кровеносными сосудами, которая на переднем полюсе глаза соединяется с роговицей. Роговица является прозрачным защитным наружным образованием, кривизна поверхности которого определяет особенности преломления света. Так, при неправильной кривизне роговицы возникает искажение зрительных изображений, называемое астигматизмом. Позади роговицы находится радужная оболочка, цвет которой зависит от пигментации составляющих ее клеток и их распределения. Между роговицей и радужной оболочкой находится передняя камера глаза, наполненная жидкостью тАФ "водянистой влагой". В центре радужной оболочки находится зрачок круглой формы, пропускающий внутрь глаза свет после его прохождения через роговицу.
Размер зрачка зависит от освещенности. Контроль за изменениями размера зрачка осуществляется автоматически нервными волокнами, заканчивающимися в мускулатуре радужной оболочки. Круговая мышца, суживающая зрачок тАФ сфинктер тАФ иннервируется парасимпатическим волокнами, мышца, расширяющая зрачок тАФ дилататор тАФ иннервируется симпатическими волокнами. Изменения диаметра зрачка меняют интенсивность светового раздражения незначительно тАФ всего в 16-17 раз (если учитывать, что диапазон интенсивности света изменяется в 16 млрд. раз). Реакция расширения зрачка до максимального диаметра тАФ 7,5 мм тАФ очень медленная: она длится около 5 минут. Максимальное сокращение диаметра зрачка до 1,8 мм достигается быстрее тАФ всего за 5 секунд. Это значит, что основная функция зрачка состоит не в регуляции интенсивности света вообще, а в том, чтобы пропускать лишь тот свет, который попадает на центральную часть хрусталика, где фокусировка наиболее точная. Сужение зрачка направлено на сохранение наиболее возможной при данных условиях освещенности глубины резкости.
Роговица и коньюктива покрыты тонкой пленкой слезной жидкости, секретируемой в слезных железах, расположенных в височной части глазницы, над глазным яблоком. Слезы защищают роговицу и коньктиву от высыхания.
Позади радужной оболочки расположены задняя камера глаза и хрусталик. Хрусталик тАФ двояковыпуклая линза, расположенная в сумке, волокна которой соединены с ресничными мышцами и наружным сосудистым слоем сетчатки. Хрусталик может становиться более плоским или более выпуклым в зависимости от расстояния между глазом и объектом. Изменение кривизны хрусталика называется аккомодацией. Внутри глаза, позади хрусталика, находится стекловидное тело. Оно представляет собой коллоидный раствор гиалуроновой кислоты во внеклеточной жидкости.
Аккомодация хрусталика иногда оказывается недостаточной, чтобы спроецировать изображение точно на сетчатку. Если расстояние между хрусталиком и сетчаткой больше, чем фокусное расстояние хрусталика, то возникает близорукость (миопия). Если сетчатка расположена слишком близко к хрусталику и фокусировка хороша только при рассматривании далеко расположенных предметов, возникает дальнозоркость (гиперметропия). Близорукость и дальнозоркость корректируются очками с вогнутыми и выпуклыми линзами соответственно.
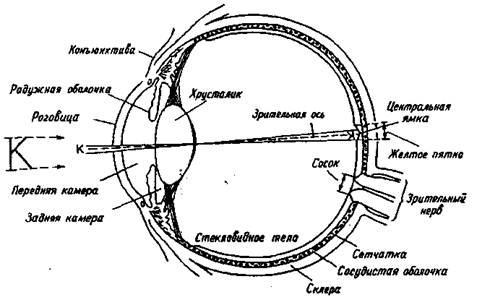
Рис. 1.1. Горизонтальный срез правого глаза
Астигматизм (результат неравномерной кривизны роговицы) плохо корректируется даже сложными линзами. Для его исправления более пригодны контактные линзы, которые, плавая в слезной жидкости над роговицей, компенсируют ее отклонения от правильной формы. Итак, оптическая система глаз обеспечивает фокусировку изображения на рецепторной поверхности сетчатки. Диоптрический аппарат, состоящий из системы линз, передает на сетчатку резко уменьшенное изображение предметов (рис. 1.1).
Сетчатка тАФ с нейроанатомической точки зрения тАФ высокоорганизованная слоистая структура, объединяющая рецепторы и нейроны (рис. 1.2). Фоторецепторные клетки тАФ палочки и колбочки тАФ расположены в пигментном слое, наиболее удаленном от хрусталика.
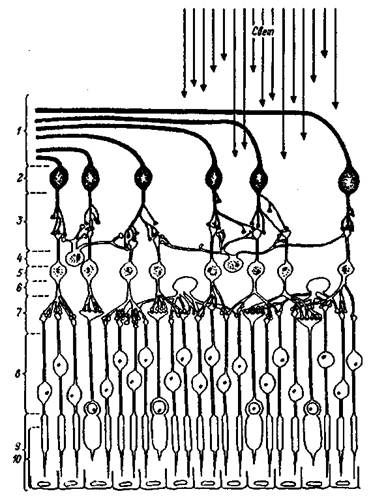
Рис. 1.2. Строение сетчатки глаза
Вверху тАФ падающий свет; 1 тАФ волокна зрительного нерва; 2 тАФ ганглиозные клетки; 3 тАФ внутренний синаптический слой; 4 тАФ амакриновые клетки; 5 тАФ биполярные клетки; 6 тАФ горизонтальные клетки; 7 тАФ наружный синаптический слой, 8 тАФ ядра рецепторов; 9 тАФ рецепторы; 10 тАФ пигментный слой эпителиальных клеток.
Они повернуты от пучка падающего света таким образом, что их светочувствительные концы спрятаны в промежутках между сильно пигментированными эпителиальными клетками. Эпителиальные пигментные клетки участвуют в метаболизме фоторецепторов и синтезе зрительных пигментов. Все нервные волокна, выходящие из сетчатки, лежат в виде переплетенного пучка на пути света, создавая препятствие на пути его попадания в рецепторы. Кроме того, в том месте, где они выходят их сетчатки по направлению к мозгу, отсутствуют светочувствительные элементы тАФ это так называемое слепое пятно. Свет, попадающий на сетчатку в области слепого пятна не воспринимается элементами сетчатки, поэтому остается "дефект" изображения, проецируемого на сетчатку. Однако, наличие слепого пятна не сказывается на целостности зрительного восприятия. Этот эффект или, точнее, дефект слепого пятна компенсируют высшие зрительные центры.
Палочки и колбочки отличаются как структурно, так и функционально. Зрительный пигмент (пурпур тАФ родопсин) тАФ содержится только в палочках. В колбочках находятся другие зрительные пигменты тАФ иодопсин, хлоролаб, эритлаб, необходимые для цветового зрения. Палочка в 500 раз более чувствительна к свету, чем колбочка, но не реагирует на свет с разной длиной волны, т.е. она не цветочувствительна. Зрительные пигменты расположены в наружном сегменты палочек и колбочек. Во внутреннем сегменте находится ядро и митохондрии, принимающие участие в энергетических процессах при действии света.
В глазу человека около 6 млн. колбочек и 120 млн. палочек тАФ всего около 130 млн. фоторецепторов. Плотность колбочек наиболее высока в центре сетчатки и падает к периферии. В центре сетчатки, в небольшом ее участке, находятся только колбочки. Этот участок называется центральной ямкой. Здесь плотность колбочек равна 150 тысячам на 1 квадратный миллиметр, поэтому в области центральной ямки острота зрения максимальна. Палочек в центре сетчатки очень мало, их больше на периферии сетчатки, но острота "периферического" зрения при хорошей освещенности невелика. В условиях сумеречного освещения преобладает периферическое зрение, а острота зрения в области центральной ямки падает. Таким образом, колбочки функционируют при ярком свете и выполняют функцию восприятия цвета, палочкой воспринимают свет и обеспечивают зрительное восприятие при слабой освещенности.
Первичный процесс зрительной рецепции тАФ фотохимическая реакция. Фотоны поглощаются молекулами зрительных пигментов. Каждая молекула пигмента поглощает один фотон (квант света) и переходит на более высокий энергетический уровень. Поглощение кванта света в фоторецепторе запускает многоступенчатый процесс распада молекул пигмента. Родопсин тАФ зрительный пигмент палочек тАФ состоит из белка (опсина) и ретиналя (альдегида витамина А1). При распаде родопсина образуются опсин и витамин А1. Иодопсин тАФ основной пигмент колбочек тАФ также состоит из опсина и ретиналя. Фотохимические процессы в палочках и колбочках сходны. Родопсин и иодопсин имеют разные спектры поглощения: максимум спектра поглощения родопсина тАФ 500 им (зелено-голубая часть), максимум спектра иодопсина тАФ 570 нм (желтая часть). Каждая палочка в сетчатке человека содержит один пигмент, каждая колбочка тАФ три разных пигмента, максимумы поглощения которых составляют примерно 425, 435 и 570 нм. Восстановление пигментов осуществляется в темноте в результате цепи химических реакций (ресинтез), протекающих с поглощением энергии. Ретиналь ресинтезируется на основе цис-изомера витамина А1, поэтому при недостатке витамина А1 в организме возникает недостаточность сумеречного зрения. Если освещение постоянно и равномерно, то фотохимический распад пигментов находится в равновесии с их ресинтезом. Этот фотохимический процесс обеспечивает светотемновую адаптацию.
При освещении фоторецептора возникает увеличение элетроотрицателъности потенциала внутри клетки по отношению к внеклеточному пространству. Это приводит к уменьшению трансмембранного тока в рецепторах. Таким образом, на свет в фоторецепторах возникает гиперполяризационный ответ. Гиперполяризация отличает зрительные рецепторы от других рецепторов, например, слуховых и вестибулярных, где возбуждение связано с деполяризацией мембраны. Амплитуда рецепторного зрительного потенциала увеличивается при увеличении интенсивности света (освещенности, относительно предыдущего состояния адаптации). Амплитуда рецепторного потенциала зависит также от длины волны света, максимум ответа палочек проявляется при длине волны максимального поглощения родопсина тАФ 500 нм, колбочек тАФ 560-570 нм.
Палочки и колбочки соединены с биполярными нейронами сетчатки, которые, в свою очередь, образуют с ганглиозными клетками синапсы, выделяющие ацетилхолин. Аксоны ганглиозных клеток сетчатки в составе зрительного нерва идут к различным мозговым структурам. Около 130 млн. фоторецепторов связаны с 1,3 млн., волокон зрительного нерва, что свидетельствует о конвергенции зрительных структур и сигналов. Только в центральной ямке каждая колбочка связана с одной биполярной клеткой, а она, в свою очередь, тАФ с одной ганглиозной. К периферии от центральной ямки на одной биполярной клетке конвергируют множество палочек и несколько колбочек, а на ганглиозной тАФ множество биполярных. Поэтому функционально такая система обеспечивает переработку первичного сигнала, повышающую вероятность его обнаружения за счет широкой конвергенции связей от периферических рецепторов к ганглиозной клетке, посылающей сигналы в мозг (рис. 1.3).
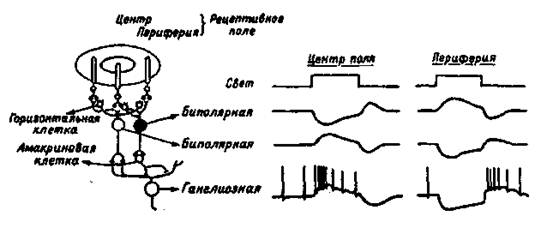
Рис. 1.3. Организация концентрических рецептивных полей биполярных и ганглиозных клеток сетчатки (слева) и схема изменений потенциала, построенная на основе внутриклеточных записей (справа).
Два тина тормозных нейронов тАФ горизонтальные и амакриновые клетки тАФ расположены в том же слое, где находятся биполярные нейроны, ограничивают распространение зрительного возбуждения внутри сетчатки. Горизонтальные и амакриновые клетки связаны с биполярными и ганглиозными горизонтальными связями, обеспечивающими латеральное торможение между соседними клеточными элементами сетчатки: горизонтальные тАФ между биполярными, амакриновые тАФ между ганглиозными.
Ганглиозные клетки сетчатки при слабой освещенности дают непрерывную импульсацию. При усилении освещенности половина клеток усиливает импульсацию, половина тАФ ослабляет. Следовательно, первые являются детекторами яркости, вторые тАФ детекторами темноты. Все ганглиозные клетки сетчатки имеют круглые рецептивные поля в отличие от неправильных нессиметричных рецептивных полей слуховых и соматических нейронов. Оптимальным стимулом для ганглиозных клеток служит либо светлое пятно, окруженное темным поясом, либо темное пятно, окруженное ярким поясом. Многие ганглиозные клетки реагируют только на изменение освещения, но не реагируют на постоянный свет. Схематическое изображение рецептивных полей и реакций нейронов сетчатки показано на рис.1.3.
Каждая ганглиозная клетка имеет свое рецептивное поле, т.е. ограниченный участок сетчатки, фоторецепторы которого связаны с данной клеткой. Реакция такой клетки на свет вне ее рецептивного поля отсутствует. Ганглиозные клетки функционально различны, их разделяют на два типа:
1) нейроны, которые возбуждаются светом, падающим на центр рецептивного поля, но затормаживаются, если свет падает на его периферию;
2) нейроны, которые затормаживаются светом в центре рецептивного поля и возбуждаются при действии света на его края.
Одновременная реакция нейронов первого и второго типов лежит в основе появления одновременного контраста за счет подчеркивания края изображения их антагонистическими рецептивными полями.
Размеры рецептивных полей, равно как и их функциональные свойства, зависят от расположения фоторецепторов данного поля на сетчатке относительно центральной ямки (эксцентриситет). Величина рецептивных полей растет от области центральной ямки к периферии сетчатки. Это является следствием структурной организации связей элементов сетчатки. Так, в середине сетчатки, в области центральной ямки, где плотность колбочек максимальная, одна колбочка через отдельную биполярную клетку соединяется с отдельной ганглиозной клеткой. Таким образом, рецептивные поля ганглиозных клеток, связанные с центром сетчатки очень узкие и не перекрываются. На периферии, где, в основном, находятся палочки, отмечаются широкие рецептивные поля: множество рецепторов связано с одной ганглиозной клеткой.
Функционально острота зрения зависит и от свойств рецептивных полей: острота зрения тАФ возможность различения двух соседних точек тАФ максимальна для узких рецептивных полей центральной ямки. В то же время слабые сигналы с периферии сетчатки выделяются зрением, благодаря взаимодействию широких перекрывающихся рецептивных полей за счет пространственной суммации раздражителей.
При освещении сетчатка генерирует электрические потенциалы, которые называют электроретинограммой. Эта суммарная электрическая реакция отражает процессы возбуждения различных нервных структур: волна возникает во внутренних сегментах фоторецепторов; волна является результатом возбуждения биполярных и амакриновых клеток сетчатки; волна связана с пигментным эпителием; волна, является реакций горизонтальных клеток сетчатки на выключение света.
Таким образом, волны электроретинограммы имеют своим источником все клеточные элементы сетчатки, кроме ганглиозных.
В сетчатке обнаружены клетки, для которых наиболее эффективны цвета-антагонисты красный и зеленый, а также клетки, для которых пару антагонистов составляют желтый и синий или зеленый и синий. Объяснение антагонистического действия цветов на ганглиозные клетки состоит в том, что из трех типов колбочек два всегда связаны с одним нейроном, а часть колбочек имеет возбудительные синапсы, другая часть тАФ тормозные.
Сетчатку, по сложности организации, часто рассматривают как часть мозга, расположенную на периферии. Здесь с фоторецепторами связаны несколько слоев нейронов, формирующих афферентный поток, который идет к подкорковым и корковым центрам зрительной системы. Горизонтальные и биполярные клетки сетчатки не генерируют потенциалов действия, основной формой их активности являются градуальные гиперполяризации и деполяризации. Ганглиозные клетки генерируют потенциалы действия, которые проводятся по их длинным аксонам, составляющим зрительный нерв.
Зрительный нерв содержит около 800 тысяч волокон ганглиозных клеток сетчатки. Зрительные нервы обоих глаз перекрещиваются в области основания черепа, где около полумиллиона волокон зрительного нерва переходят на противоположную сторону. Остальные 300 тысяч волокон вместе с перекрещенными аксонами второго зрительного нерва образуют зрительный тракт.
Нервные волокна зрительного тракта подходят к четырем структурам мозга: ядрам верхних бугров четверохолмия тАФ средний мозг; ядрам латерального коленчатого тела тАФ таламус; супрахиазмальным ядрам гипоталамуса; к глазодвигательным нервам.
Ядра верхних бугров четверохолмия и латерального коленчатого тела являются конечными пунктами двух параллельных путей от ганглиозных клеток сетчатки: одна ветвь аксона ганглиозной клетки идет в латеральное коленчатое тело, другая тАФ в верхнее двухолмие. Обе ветви сохраняют упорядоченную проекцию сетчатки. От переднего двухолмия после переключения сигналы идут к крупному ядру таламуса тАФ подушке.
Аксоны клеток латерального коленчатого тела, проходящие в составе зрительной радиации, проецируются к клеткам первичной зрительной коры (поле 17 или стриарная кора). Проекция зрительной ямки сетчатки тАФ зоны максимальной остроты зрения тАФ в 35 раз больше проекции участка такого же размера на периферии сетчатки. Клетки поля 17 (стриарной коры) связаны с полями 18 и 19 (престриарная кора), так называемыми вторичными зрительными зонами. От этих зон идут проекции к подушке таламуса, куда поступает информация от верхних бугров четверохолмия. Кроме того, зрительные пути прослеживаются к лобной коре, они примыкают к ассоциативной коре.
Клетки латерального коленчатого тела, получающие основную афферентацию от сетчатки, имеют простые концентрические рецептивные поля, как и ганглиозные клетки. Здесь проявляется бинокулярное взаимодействие: волокна от обоих глаз распределены топографически правильно, послойно. В то же время небольшая часть клеток латерального коленчатого тела активируется от обоих зрительных нервов.
Нейроны зрительной коры уже имеют не концентрические, а почти прямоугольные зрительные поля, некоторые из нейронов реагируют на определенную ориентацию (наклон) полосы тАФ светлой или темной (рис. 1.4).
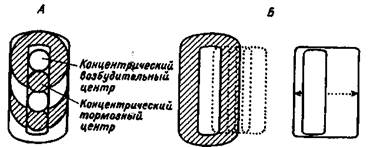
Рис. 1.4. Концентрические рецептивные поля в сетчатке и подкорковых зрительных центрах (А), прямоугольные и сложные рецептивные поля в зрительной коре (Б).
В зрительной коре существуют два функционально различных типа клеток: простые и сложные. Простые клетки имеют рецептивное поле, состоящее из возбудительной и тормозной зоны, которые можно предсказать на основе исследования реакции клетки на маленькое световое пятно. Структуру рецептивного поля сложной клетки невозможно установить сканированием светового пятнышка. Они служат "детекторами" угла, наклона или движения линий в поле зрения. В коре уже совершенно отчетлива бинокулярная конвергенция: в одной точке представлены симметричные поля зрения тАФ справа и слева.
Близко расположенные клетки зрительной коры "видят" только небольшую часть поля зрения. Лежащие друг под другом нейроны одной колонки коры реагируют на один и тот же стимул, оптимальный по ориентации, наклону и направлению движения. В одной колонке могут располагаться как простые, так и сложные клетки. Простые клетки найдены в III и IV слоях, где заканчиваются таламические волокна. Сложные клетки расположены в более поверхностных слоях коры 17 поля. В полях 18 и 19 зрительной коры простые клетки являются исключением, здесь расположены сложные и сверхсложные клетки. Последние реагируют, например, только на стимулы определенной ширины, длины и ориентации.
Итак, от уровня к уровню зрительной системы происходит усложнение рецептивных полей нейронов. Все рецептивные поля организованы в виде возбудительных и тормозных зон. Концентрические рецептивные поля, характерные для сетчатки и латерального коленчатого тела, уже не встречаются в коре. В зрительной системе, как и в других сенсорных системах, чем выше синаптический уровень, тем строже ограничены функции отдельных нейронов тАФ детекторов свойств.
Для успешной работы системы распознавания зрительных образов очень важны движения глаз. Известно, что глаз человека приводится в движение шестью наружными мышцами. Относительно координат головы глаза двигаются горизонтально, вертикально и вокруг своей оси. Если оба глаза двигаются в одном направлении, такие движения называются содружественными. При переводе взгляда с ближней точки на дальнюю осуществляются дивергентные движения. При наклоне головы в сторону наблюдаются небольшие вращательные движения глаз.
При взгляде на любой предмет глаза двигаются от одной точки фиксации к другой быстрыми скачками тАФ саккадами. Длительность саккад от 10 до 80 мс, длительность периодов фиксации 150-300 мс. Медленные движения глаз реализуются при слежении за движущимися объектами тАФ следящие движения.
Движения глаз управляются центрами, которые находятся в области ретикулярной формации мозга и среднего мозга, в верхних буграх четверохолмия и в претектальной области. Все эти подкорковые центры координируются сигналами из зрительной, теменной и лобной коры, ответственными за программирование движений тела и оценки его положения в пространстве. Для наиболее тонкой регуляции глазодвигательных функций весьма существенны влияний мозжечка, сравнивающего тонический и фазный компоненты движения при ориентации и пространстве.
В процессе зрительного восприятия, особенно при слежении за движущимся объектом, возникает оптический нистагм, вызываемый движущимися оптическими стимулами и состоящий из чередовании саккад и медленные следящих движений. Движения глаз имеют огромное значение для восприятия: при неподвижном глазном яблоке восприятие изображения пропадает в связи с разложением пигмента и адаптацией фоторецепторов.
Координированные движения глаз обеспечивают объединение информации, идущей от обоих глаз в центры мозга. Особое значение для восприятия и координации движений играют нейроны переднего двухолмия. Они организованы в колонки, которые воспринимают сигналы, поступающие от одних и тех же участков полей зрения: активность нейронов мою отдела мозга, на которых конвергирует импульсация от правого и левого глаза, является пусковым механизмом для глазодвигательных нейронов. На высших уровнях зрительной системы параллельно функционируют две системы анализа: одна определяет место предмета и пространстве, другая описывает его признаки. Конечные результаты параллельных процессов интегрируются и возникает законченный зрительный образ внешнего предметного мира.
2 СОМАТОВИiЕРАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
2.1 Психофизика кожной механорецепции
Пороги субъективная интенсивность тактильных стимулов
Абсолютный порог для тактильного стимула можно измерить калиброванными волосками по методу, который ввел в психофизику в XIX в. немецкий физиолог Макс фон Фрей (Max von Frey). В современных стимуляторах высокоточные движения раздражающего стерженька обеспечиваются электромагнитом. Стерженек обеспечивает воздействие градуированной интенсивности с заданной временной характеристикой, например одиночные импульсы или ритмичные нажимы с синусоидально изменяющейся интенсивностью (вибрации).
Пороги восприятия, определенные волосками Фрея и вибрациями (200 Гц), генерируемыми электромагнитным стимулятором, приведены для разных частей тела в табл. 2.1.
Таблица 2.1. Пороги восприятия механической стимуляции кожи человека
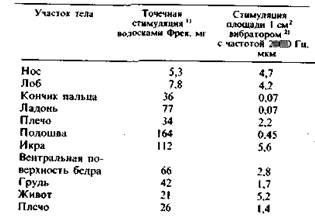
Кончики пальцев воспринимают вибрацию амплитудой менее 1 мкм. Относительная чувствительность разных частей тела весьма неодинакова. Кроме того, разные по своей природе стимулы демонстрируют неодинаковую чувствительность одного и того же места (табл. 2.1). Это объясняется тем, что околопороговая вибрация и волоски Фрея стимулируют разные типы механорецепторов.
Пространственное разрешение тактильных стимулов
Для получения меры пространственного paзрешения пригодны различные тесты по распознаванию пространственных деталей тактильных стимулов. В одном из часто применяемых тестов используя циркуль, двумя кончиками которого одновременно прикасаются к коже (рис. 2.1) для определения двухточечного порога.
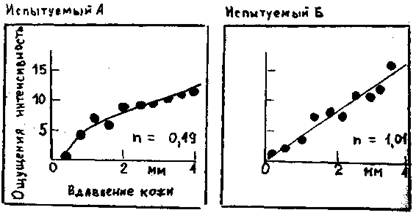
Рис. 2.1. Психофизические опыты для оценки интенсивности стимула. Испытуемый выражает ощущаемую им силу давления на ладонь (здесь тАУ длительностью 1с.) числом на произвольно выбранной шкале (ось ординат). Объективную интенсивность стимула (ось обсцисс) меняли в случайном порядке. Кривые тАУ степенные функции, соответствующие полученным данным с показателем степени n, отмеченным графиках.
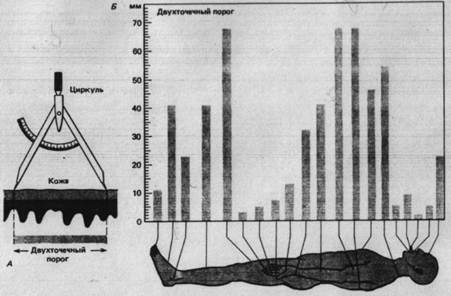
Рис. 2.2. Двухточечный порог кожи. А. Кончики циркуля, разведенные на разные расстояния, неоднократно прикасаются к коже. Двухточечный порог - наименьшее расстояние, при котором испытуемый может определить, чтo стимулируются две точки, а не одна. Б. Изменчивость двухточечного порога кожи в зависимости oт участка тела человека.
Испытуемый должен, не глядя, ответить, в одной или в двух точках до него дотронулись. Двухточечный порог на разных частях тела неодинаков (рис. 2.2 Б). Это обусловлено особенности иннервации разных областей кожи и связанным с ней характером соединения афферентных волокон в ЦНС.
2.2 Кожные механорецепторы
Исследования на человеке и животных привели к хорошему пониманию физиологической функции гистологического строения механорецепторов кожи и обнаружили между ними корреляцию. Кожа млекопитающих содержит несколько основных типов механорецепторов.
Типы низкопороговых механорецепторов кожи
Нейрофизиологические опыты показали, что лишенная волос кожа крыс, кошек, обезьян и людей одержит три основных типа механорецепторов с афферентами группы II (А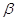 ). Их называют рецепторами МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном воздействии механического стимула (давления; например, давления веса тела на подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток импульсов. БА тАУ это быстро адаптирующийся рецептор, реагирующий только на механические стимулы, которые изменяются во времени. ТП означает тельца Пачини, гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся механорецепторы.
). Их называют рецепторами МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном воздействии механического стимула (давления; например, давления веса тела на подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток импульсов. БА тАУ это быстро адаптирующийся рецептор, реагирующий только на механические стимулы, которые изменяются во времени. ТП означает тельца Пачини, гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся механорецепторы.
Рецептивные поля и плотность иннервации механорецепторов
Область, в которой стимул данной интенсивности способен возбудить механочувствительное афферентное волокно, называется его рецептивным полем. Оно приблизительно соответствует анатомической протяженности всех окончаний этого волокна. Например, один афферент связан с двумя-тремя тактильными тельцами оволосненной кожи, а все 30-50 дисков Меркеля такого тельца иннервируются одной коллатералью. Концевые разветвления гораздо протяженнее у рецепторов волосяных фолликулов: каждый афферент может отходить от многих фолликулов, и каждый фолликул иннервируется несколькими афферентными волокнами.
Какие свойства рецепторов определяют пространственное разрешение, и
Вместе с этим смотрят:
РЖсторiя виникнення та розвитку масажу
Аборты
Аденовирусная инфекция
Азотные и кислородные ванны, нафталановая нефть
Акушерська операцiя - накладання акушерських щипцiв