Структура центральной нервной системы и общие принципы её функционирования
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ УКРАИНЫ
ЛУБЕНСКОЕ МЕДИЦИНСКОЕ УЧИЛИЩЕ
РЕФЕРАТ
iИЗИОЛОГИИ
НА ТЕМУ:
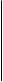 Структура центральной нервной системы и общие принципы её функционирования
Структура центральной нервной системы и общие принципы её функционирования
Выполнила:
студентка группы Ф-31
Николаева Юлия
Лубны 2009
Центральная нервная система включает головной и спинной мозг, выполняющие в организме человека и животных сложнейшие функции.
Функции центральной нервной системы. 1. Центральная нервная система обеспечивает взаимную связь отдельных органов и систем, согласует и объединяет их функции. Благодаря этому организм работает как единое целое. Точность контроля за работой внутренних органов достигается существованием двусторонней круговой связи между центральной нервной системой и периферическими органами.
Центральная нервная система осуществляет связь организма с внешней средой, обеспечивает индивидуальное приспособление к внешней среде тАФ поведение человека и животных.
Головной мозг является органом психической деятельности. В результате поступления нервных импульсов в клетки коры большого мозга возникают ощущения и на их основе проявляются специфические качества высокоорганизованной материи тАФ процессы сознания и мышления.
Центральная нервная система состоит из серого и белого вещества. Серое вещество представляет собой скопление нервных клеток, белое вещество состоит из их отростков. Кроме нервных клеток, в центральной нервной системе имеется нейроглия. Она со всех сторон окружает нейроны.
Структура и функция нейронов (нервные клетки). В каждом нейроне различают тело и отростки тАФ аксон и дендриты.
Аксои тАФ длинный отросток тАФ проводит возбуждение от тела нервной клетки к другим нейронам или к периферическим органам; дендритытАФкороткие, сильно ветвящиеся отростки тАФ осуществляют связь между отдельными нервными клетками.
Тело нервной клетки и ее отростки покрыты мембраной, избирательно проницаемой в состоянии покоя, главным образом для ионов калия, а при возбуждении тАФ преимущественно для ионов натрия. В условиях покоя мембранный потенциал различных нервных клеток обычно равен 50тАФ70 мВ. При возбуждении возникающий потенциал действия составляет 80тАФПО мВ.
Внутри нейрона находится желеобразное вещество тАФ нейроплазма. Тела нервных клеток выполняют трофическую функцию по отношению к отросткам, то есть регулируют их обмен веществ.
Нервная клетка возбуждается нервными импульсами, поступающими с периферии от рецепторов по центростремительным нервным путям или от других нейронов. Нервные клетки могут активироваться также под влиянием гуморальных воздействий. Примером являются клетки дыхательного центра, которые возбуждаются углекислым газом.
Классификация нейронов. В зависимости от выполняемой функции нейроны делятся на три основные группы: 1) воспринимающие, чувствительные, или рецептерные; 2) исполнительные, или эф-фекторные; 3) контактные, или промежуточные (вставочные).
Воспринимающие нейроны несут нервные импульсы в центральную нервную систему к рефлекторному центру. Отростки этих нервных клеток называют афферентными, или центростремительными, волокнами. Э фекторные тАФ двигательные или секреторные нейроны тАФпередают нервные импульсы от центральной нервной системы по эфферентным или центробежным волокнам.
Различным органам, изменяя их состояние и деятельность. Контактные нейроны осуществляют связь между различными нейронами.
Многие клетки б центральной нервной системе обладают автоматией. В этих клетках могут возникать нервные импульсы даже в отсутствие внешних раздражений, под влиянием продуктов обмена веществ (например, нейроны дыхательного центра).
Синапсы. В центральной нервной системе нервные клетки связаны друг с другом посредством синапсов. Синапс тАФ место контакта двух нейронов.
Одно нервное волокно может образовывать до 10 000 синапсов на многих нервных клетках.
Синапсы центральных нейронов, так же как и периферических нейронов, состоят из нервного окончания (терминали), покрытого пресинаптической мембраной, синаптической щели и постсинаптической мембраны, находящейся на теле или дендритах нейрона, которым передаются нервные импульсы.
В нервных окончаниях вырабатываются и накапливаются особые химические вещества, участвующие в передаче возбуждения через синапс. Эти вещества получили название медиаторов.
В центральной нервной системе различают возбуждающие и тормозные синапсы.
В возбуждающих синапсах под влиянием нервных импульсов освобождается возбуждающий медиатор (ацетилхолин, норадреналин, глутамат, серотонин), который через синаптическую щель поступает к постсинаптической мембране и вызывает кратковременное повышение ее проницаемости для ионов натрия и возникновения деполяризации. Когда деполяризация достигает определенного (критического) уровня, возникает распространяющееся возбуждение тАФ потенциал действия.
В тормозных синапсах выделяются особые тормозные медиаторы (ГАМК тАФ гамма-аминомасляная кислота и др.). Они изменяют проницаемость постсинаптической мембраны по отношению к ионам калия или хлора. В результате повышается уровень мембранного потенциала тАФ явление гиперполяризации, что препятствует дальнейшему распространению возбуждения.
Рефлекс. Рефлекторная дуга. Виды рефлексов.
Основной формой нервной деятельности является рефлекс. Рефлекс тАФ причинно обусловленная реакция организма на изменения внешней или внутренней среды, осуществляемая при участии центральной нервной системы в ответ на раздражение рецепторов. Так происходит возникновение, изменение или прекращение какой-либо деятельности организма.
Понятия о рефлекторной дуге подробно рассмотрены в главе I.
Рефлекторные дуги могут быть простыми и сложными. Простая рефлекторная дуга состоит из двух нейронов тАФ воспринимающего и эффекторного, между которыми имеется один синапс. Схема такой двухнейронной рефлекторной дуги приведена на рис. 67.
Примером простой рефлекторной дуги являются рефлекторные дуги сухожильных рефлексов, например, рефлекторная дуга коленного рефлекса.
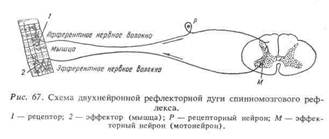
Рефлекторные дуга большинства рефлексов включают не два, а большее количество нейронов: рецепторный, один или несколько вставочных и эффекторный. Такие рефлекторные дуги называются сложными, многонейронными. Схема сложной (трехнейронной) рефлекторной дуги приведена на рис. 68.
В настоящее время установлено, что во время ответной реакции эффектора возбуждаются многочисленные нервные окончания, имеющиеся в рабочем органе. Нервные импульсы теперь уже от эффектора вновь поступают в центральную нервную систему и информируют ее о правильности ответа рабочего органа. Таким образом, рефлекторные дуги являются не разомкнутыми, а кольцевыми образованиями.
Рефлексы отличаются большим многообразием. Их можно классифицировать по ряду признаков: 1) по биологическому значению (пищевые, оборонительные, половые); 2) в зависимости от вида раздражаемых рецепторов: экстероцептивные, интероцептивные и проприоцептивные;3) по характеру ответной реакции: двигательные или моторные (исполнительный орган тАФ мышца), секреторные (эффектор тАФ железа), сосудодвигательные (сужение или расширение кровеносных сосудов).
Все рефлексы целостного организма могут быть разделены на две большие группы: безусловные и условные. Различия между ними будут разобраны в главе XI.
От рецепторов нервные импульсы по афферентным путям поступают в нервные центры. Следует различать анатомическое и физиологическое понимание нервного центра.
Нервный центр с анатомической точки зрения тАФ совокупность нейронов, расположенных в определенном отделе центральной нервной системы. За счет работы такого нервного центра осуществляется несложная рефлекторная деятельность, например, коленный рефлекс. Нервный центр этого рефлекса располагается в поясничном отделе спинного мозга (IIтАФIV сегменты).
Нервный центр с физиологической точки зрения тАФ сложное функциональное объединение нескольких анатомических нервных центров, расположенных на разных уровнях центральной нервной системы и обусловливающих за счет своей активности сложнейшие рефлекторные акты. Например, в осуществлении пищевых реакций участвуют многие органы (железы, мышцы, кровеносные и лимфатические сосуды и т. д.). Деятельность этих органов регулируется нервными импульсами, поступающими из нервных центров, располагающихся в различных отделах центральной нервной системы. А. А. Ухтомский эти функциональные объединения назвал ВлсозвездиямиВ» нервных центров.
Физиологические свойства нервных центров. Нервные центры обладают рядом характерных функциональных свойств, зависящих от наличия синапсов и большого количества нейронов, входящих в их состав. Основными свойствами нервных центров являются: 1) одностороннее проведение возбуждения; 2) задержка проведения возбуждения; 3) суммация возбуждений; 4) трансформация ритма возбуждений; 5) рефлекторное последействие; 6) быстрая утомляемость.
Одностороннее проведение возбуждения в центральной нервной системе обусловлено наличием в нервных центрах синапсов, в которых передача возбуждения возможна только в одном направлении тАФ от нервного окончания, выделяющего медиатор, к постсинаптической мембране.
Задержка проведения возбуждения в нервных центрах также связана с наличием большого количества синапсов. На выделение медиатора, его диффузию через синаптическую щель, возбуждение постсинаптической мембраны требуется больше времени, чем на распространение возбуждения по нервному волокну.
Суммация возбуждений в нервных центрах возникает или при нанесении слабых, но повторяющихся (ритмичных) раздражений, или при одновременном действии нескольких подпороговых раздражений. Механизм этого явления связан с накоплением медиатора на постсинаптической мембране и повышением возбудимости клеток нервного центра. Примером суммации возбуждения может служить рефлекс чихания. Этот рефлекс возникает только при длительном раздражении рецепторов слизистой оболочки носа. Впервые явление суммации возбуждений в нервных центрах было описано И. М. Сеченовым в 1863 г.

Трансформация ритма возбуждений заключается в том, что центральная нервная система на любой ритм раздражения, даже медленный, отвечает залпом импульсов. Частота возбуждений, поступающих из нервных центров на периферию к рабочему органу, колеблется от 50 до 200 в секунду, Этой особенностью центральной нервной системы объясняется то, что все сокращения скелетных мышц в организме являются титаническими.
Рефлекторные акты заканчиваются не одновременно с прекращением вызвавшего их раздражения, а через некоторый, иногда сравнительно длительный, период. Это явление получило название рефлекторного последействия.
Установлены два механизма, обусловливающие последействие. Первый связан с тем, что возбуждение в нервных клетках исчезает не сразу после прекращения раздражения.В течение некоторого времени (сотые доли секунды) нервные клетки продолжают давать ритмические разряды импульсов. Этот механизм может обусловить лишь сравнительно кратковременное последействие. Второй механизм является результатом циркуляции нервных импульсов по замкнутым нейронным цепям нервного центра и обеспечивает более длительное последействие. На рис. 69 показана такая замкнутая цепь нейронов.
Возбуждение одного из нейронов передается на другой, а по ответвлениям его аксона вновь возвращается к первой нервной клетке и т. д. Циркуляция нервных импульсов в нервном центре будет продолжаться до тех пор, пока не наступит утомление одного из синапсов или же активность нейронов не будет приостановлена приходом тормозных импульсов.
Нервные центры легко утомляемы в отличие от нервных волокон. При продолжительном раздражении афферентных нервных волокон утомление нервного центра проявляется постепенным снижением, а затем и полным прекращением рефлекторного ответа.
Эта особенность нервных центров доказывается следующим образом. После прекращения мышечного сокращения в ответ на раздражение афферентных нервов начинают раздражать эфферентные волокна, иннервирующие мышцу. В этом случае мышца вновь сокращается. Следовательно, утомление развилось не в афферентных путях, а в нервном центре.

Рефлекторный тонус нервных центров. В состоянии относительного покоя, без нанесения дополнительных раздражении, из нервных центров на периферию к соответствующим органам и тканям поступают разряды нервных импульсов, В покое частота разрядов и количество одновременно работающих нейронов очень небольшие. Редкие импульсы, непрерывно поступающие из нервных центров, обусловливают тонус (умеренное напряжение) скелетных мышц, гладких мышц кишечника и сосудов. Такое постоянное возбуждение нервных центров носит название тонуса нервных центров. Он поддерживается афферентными импульсами, непрерывно поступающими от рецепторов (особенно проприорецепторов), и различными гуморальными влияниями (гормоны, С02 и др.).
Торможение (как и возбуждение) тАФ активный процесс. Торможение возникает в результате сложных физико-химических изменений в тканях, но внешне этот процесс проявляется ослаблением функции какого-либо органа.
В 1862 г. были проведены классические опыты основоположником русской физиологии И. М. Сеченовым, получившие название Влцентральное торможениеВ». На зрительные бугры лягушки, отделенные от больших полушарий головного мозга, И. М. Сеченов помещал кристаллик хлорида натрия (поваренной соли) и наблюдал при этом торможение спинномозговых рефлексов. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась (рис. 70).
Результаты этого опыта позволили И. М. Сеченову заключить, что в центральной нервной системе наряду с процессом возбуждения развивается и процесс торможения, способный угнетать рефлекторные акты организма.
В настоящее время принято выделять две формы торможения: первичное и вторичное.
Для возникновения первичного торможения необходимо наличие специальных тормозных структур (тормозных нейронов и тормозных синапсов). Торможение в этом случае возникает первично без предшествующего возбуждения.
Примерами первичного торможения могут служить пре- и постсинаптическое торможение. Пресинаптическое торможение развивается в аксо-аксональных синапсах, образованных на пресинаптических окончаниях нейрона. В основе пресинаптического торможения лежит развитие медленной и длительной деполяризации пресинаптического окончания, что приводит к уменьшению или блокаде дальнейшего проведения возбуждения. Постсинаптическое торможение связано с гиперполяризацией постсинаптической мембраны под влиянием медиаторов, которые выделяются при возбуждении тормозных нейронов.
Первичное торможение играет большую роль в ограничении, поступления нервных импульсов к эффекторным нейронам, что имеет существенное значение в координации работы различных отделов центральной нервной системы.
Для возникновения вторичного торможения не требуется специальных тормозных структур. Оно развивается в результате изменения функциональной активности обычных возбудимых нейронов.
Значение процесса торможения. Торможение наряду с возбуждением принимает активное участие в приспособлении организма к окружающей среде. Торможение играет важную роль в формировании условных рефлексов: освобождает центральную нервную систему от переработки менее существенной информации; обеспечивает координацию рефлекторных реакций, в частности, двигательного акта. Торможение ограничивает распространение возбуждения на другие нервные структуры, предотвращая нарушение их нормального функционирования, то есть торможение выполняет охранительную функцию, защищая нервные центры от утомления и истощения.
Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации.
Явление координации играет важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров.
За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за . счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т. д.).
Установлен ряд общих закономерностей тАФ принципов координации: 1) принцип конвергенции; 2) принцип иррадиации возбуждения; 3) принцип реципрокности; 4) принцип последовательной смены возбуждения торможением и торможения возбуждением; 5) феномен ВлотдачиВ»; 6) цепные и ритмические рефлексы; 7) принцип общего конечного пути; 8) принцип обратной связи; 9) принцип доминанты.
Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, приходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эффекторным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эффекторных. Поэтому афферентные нейроны образуют на телах и дендритах эффекторных и вставочных нейронов многочисленные синапсы.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.
Принцип реципрокности (сопряженности). Это явление было изучено И. М. Сеченовым, Н. Е. Введенским, Шеррингтоном. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам антагонистов мышц-сгибателей и разгибателей конечностей. Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное). Если раздражать у спинального животного (кошка) кожу конечности, отмечается сгибательный рефлекс данной конечности, а иа противоположной стороне в это время наблюдается рефлекс разгибания. Описанные явления связаны с тем, что при возбуждении центра сгибания одной конечности происходит реципрокное торможение центра разгибания этой же конечности. На симметричной стороне имеются обратные взаимоотношения: возбужден центр разгибателей и заторможен центр сгибателей. Только при такой взаимосочетанной (реципрокной) иннервации возможна ходьба.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и прочее,
Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы, Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эффекторных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям. Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эффекторные нейроны). Общими путями могут быть не только конечные эффекторные нейроны, но и вставочные.
Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов) , получили название вторичных афферентных импульсов или Влобратной с в я з иВ».
Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные тАФ их угнетению.
Принцип доминанты был сформулирован А. А, Ухтомским. Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта тАФ временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами: 1) повышенной возбудимостью; 2) стойкостью возбуждения; 3) способностью к суммированию возбуждения; 4) инерцией тАФ доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы от других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
Использованная литература
1. Физиология / Под ред. С.А. Георгиевой. тАУ 2-е изд. тАУ Ф48 М.: Медецина, 1986. тАУ 400 с.
Вместе с этим смотрят:
РЖсторiя виникнення та розвитку масажу
Аборты
Аденовирусная инфекция
Азотные и кислородные ванны, нафталановая нефть
Акушерська операцiя - накладання акушерських щипцiв