Биология белого медведя
Министерство Сельского Хозяйства Российской Федерации
Вятская Государственная Сельскохозяйственная Академия
Биологический факультет специальность охотоведение
Кафедра Биологии промысловых зверей и птиц
Курсовая работа:
по дисциплине: ВлБиологии зверейВ»
на тему: ВлБиология белого медведяВ»
Выполнил:
ст-т 2го курса
Кашин А.В.
Проверил: д.б.н.,
профессор
Машкин В.И.
Киров 2008
ВВЕДЕНИЕ
Белый медведь, вечный скиталец среди льдов Северного Ледовитого океана, так же неотделим от арктического пейзажа, как и сами льды. Крупный, энергичный, очень сильный и в то же время миролюбивый, этот зверь - великолепная живая эмблема Арктики.
Белый медведь изучен слабо. Это объясняется тем, что он обитает в ненаселенной человеком и труднодоступной для исследователей области земного шара. Если учесть препятствия, которые встречает в Арктике ученый: дрейф льдов, хаотические нагромождения торосов, бесчисленные трещины и разводья между ледяными полями, мороз и метели, полярную ночь, длящуюся почти половину года, - станет ясно, что вести прямые наблюдения за белым медведем в природе трудно, а часто и невозможно. По этой причине скудными остаются сведения о его морфологии и физиологии, о многих особенностях биологии, в том числе о поведении и характере миграций.
Среди специалистов нет единого мнения о том, существуют ли географически обособленные группы белых медведей. И если такие группы существуют, то насколько они самостоятельны, как распределяются в пределах Арктики. Почти неизвестны состав популяций, величина годового прироста и причины смертности этого животного. Крайне слабо исследованы его приспособления к обитанию в суровых арктических условиях - низких температур и длительных голодовок.
В последнее время внимание зоологов привлекает проблема ориентации живых организмов - внутреннего механизма, помогающего им определять направление и местонахождение. Установлено, что птицы ориентируются в пространстве благодаря чувству времени, а также по положению Солнца и звезд. По протяженности миграций белый медведь, не уступает птицам, однако в пределах его ареала Солнце по нескольку месяцев не показывается над горизонтом. Можно предположить, что механизм его ориентации существенно отличается от механизма ориентации птицы. Это животное представляет интересную Влживую модельВ», использование которой перспективно в решении важных общебиологических проблем.
Белый медведь привлекает к себе большое вниманиеи как редкий вид мировой фауны. Международный союз охраны природы и природных ресурсов включил его в ВлКрасную книгуВ» - список редких видов животных земного шара. При этой организации создана специальная paбочая группа по координации исследований и охране белого медведя.
В 1974 г. между СССР, США, Норвегией, Канадой и Данией было заключено специальное соглашение о международной охране белого медведя. В последние годы в этих странах исследования, связанные с изучением белого медведя, упорядочением его использования и охран приняли особенно широкий размах.
Цель этой работы тАУ ознакомление с биологией белого медведя.
1. СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
Тип: Хордовые_ Chordata
Подтип: Позвоночные_ Vertebrata
Класс: Млекопитающие_ Mammalia
Инфракласс: Плацентарные_ Eutheria
Отряд: Хищные_Carnivora Bowdich, 1821
Семейство: Медвежьи_Ursidae Gray, 1825
Род: Медведи_ Ursus Linnaeus, 1758
Вид: Медведь белый_ Ursus maritimus Phipps, 1774
(http://" onclick="return false">
2. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
2.1 Внешний вид
Весь облик белого медведя, как и его образ жизни, отражает влияние природных условий, в которых обитает этот зверь. Белый медведь внешне заметно отличается от бурого. У него удлиненное и более узкое, особенно передней части, туловище, длинная подвижная шея, относительно небольшая голова с нешироким лбом, спрямленнымпрофилем и короткими округлыми ушами, слабо выдающимися из волосяного покрова. Неопушенная часть носа и губы - черные с сизоватым оттенком - менее подвижны, чем у бурого медведя. Хвост очень короткий, скрыт в волосяном покрове. Телосложение животного зависит от упитанности: более высокими (и подвижными) оказываются малоупитанные особи.
Ступни лап, особенно передних, у белого медведя массивные, широкие, с густоопушенными нижними поверхностями, благодаря чему площадь голых участков на них (подушки пальцев и мозоли) меньше, чем у бурого медведя. Когти темно-бурые, очень высокие и массивные, более крупные на передних лапах (длина когтя по изгибу на третьем пальце передней лапы достигает 6,5- 7 см, задней - 5-5,5 см), слабоизогнутые, острые. Роговое вещество футляра когтя исключительно прочно.
Белого медведя, очевидно, можно считать наиболее, крупным из ныне живущих наземных хищников. Длина тела (без хвоста) взрослых самцов 200-250 см (редко 285-302 см), самок- 160-250 см; высота в плечах 130- 140 см (в виде исключения - 150 см); длина хвоста (с концевыми волосами) 20-22 см.
Вес белых медведей меняется в зависимости от степени упитанности. В отдельных случаях вес самца достигает 800-1000 кг. Вес взрослых самцов, обитающих в Гренландии, составляет около 450 кг (хорошо упитанные особи весят до 500 кг), самок - 350-380 кг. Осенью при мечении на Шпицбергене самцы весили 350-400, самки - 200-250 кг. На о-ве Врангеля взрослые медведицы ранней весной при их мечении весили от 178 до 300 кг.
Мех взрослых белых медведей зимой по сравнению с мехом бурых медведей более густой, короткий, жесткий, с обилием остевых направляющих и промежуточных волос и с густым ненамокающим подшерстком: в области плеч 5-6 см, на спине и крупе 8-10, на нижней части боков и брюхе 13-15 см. Летний переходный мех более короток - около 7 см.
Окраска меха изменяется от чисто-белой и желтоватой, до соломенно-желтой, серой или почти бурой. В какой-то мере окраска меха зависит от сезона года, возраста и упитанности зверя, характера его питания. Чисто-белый мех у медведей чаще бывает осенью (по окончании линьки) или зимой. Желтоватый или золотистый мех свойствен им в конце лета, но упитанным особям - уже в начале лета и даже весной. По наблюдениям в зоопарках, желтоватый оттенок меха появляется у животных, регулярно получающих с кормом тюлений жир. Серый или бурый цвет меха приобретают звери, долгое время живущие на суше, особенно не покрытой снегом. Среди полярников распространено мнение, что у медведей, живущих среди сплоченных льдов и не имеющих доступа к воде, мех белее, чем у особей, часто плавающих или вообще проводящих много времени у воды. Мех медвежат белее, чем у| взрослых.
Половых различий в окраске меха нет. При выделки шкур, особенно после их отбеливания на солнце, желтые или серые тона меха исчезают.
Кожный покров белого и бурого медведей весьма сходен. У белого медведя он несколько толще, чем у бурого.
2.2 Череп, зубная формула
Череп у белого медведя уже, чем у бурого, со спрямленным верхним профилем, вытянут за счет удлиненной мозговой части, упрощен и низок в области лба. Для нее характерна также более короткая и облегченная нижняя челюсть. Зубной ряд укорочен; коренные зубы мельче и уже, чем у бурого медведя, имеют остробугорчатую жевательную поверхность, однако клыки у белого медведя мощнее и крупнее. Характерно, что у большинства даже старых особей белых медведей зубы оказываются менее стертыми, кариозные явления, как и патологические изменения нижней челюсти, выражены реже, чем у бурых медведей
Длина черепа у взрослых самцов достигает 353--412, у самок - 311-380 мм. При продольном распиле черепа у белого медведя выявляется меньший, чем у бурого, объем носовых и обонятельных раковин, малые объем и протяженность лобных пазух. Половая изменчивость выражена в размерах и строении черепа. Череп самки легче черепа самца, стреловидный гребень на нем короче и слабее, лоб уже, надглазничные отростки сближены, скуловые дуги слабее.
Зубная формула: i-3/3; c-1/1; p-2-4/-2-4; m-2/3 =34-42
2.3 Морфология
У белого медведя мощная мускулатура. Особенно сильно развита мускулатура задних конечностей и шеи.
Подкожные жировые накопления белого медведя могут быть значительными: на огузке слой жира достигает 11-12 см, между отдельными слоями мышц - 4 см. У очень упитанных зверей обильные жировые накопления толщиной до 3 см образуются также на внутренних органах. Особенно сильно жиреют беременные медведицы перед залеганием в берлоги. По нашим наблюдениям, на о-ве Врангеля и Земле Франца-Иосифа даже весной, во время выхода их из берлог жир покрывает тело животных почти сплошным слоем, и толщина его может достигать 5 см на спине, 10 на огузке и 3-5 см
на подошвах. Распределение подкожного жира у медведицы перед выходом ее из берлоги показано на рис. 2.
Бодрствующие животные обычно бывают менее упитанны, чем особи, лежащие в берлогах или готовящиеся к залеганию. Старые медведи обоего пола, как правило, слабо упитаны. Общий вес подкожного жира у взрослого белого медведя в отдельных случаях превышает 100 кг и может достигать 40% общего веса животного. Благодаря обильным жировым отложениям удельный вес тела белого медведя составляет менее единицы (туши убитых животных не тонут).
Жир белого медведя - как подкожный, так и внутренний - слегка желтого цвета, с характерным рыбным запахом. При температуре выше 0В°С жир жидкий, при отрицательных температурах - начинает переходить в твердое состояние. Жир белого медведя имеет высокие йодные числа и низкие числа омыления, что говорит о присутствии в его составе большого количества глицеридов высокомолекулярных непредельных и высоконепредельных жирных кислот и свидетельствует о высокой химической активности и способности легко вовлекаться в обменные процессы. Пищеварительный тракт белого медведя имеет черты плотоядности: у него укорочена, например, длина кишечника. О том же свидетельствует и сравнительно плохая способность зверя к перевариванию крахмала. Велика вместимость желудка белого медведя (свыше 70 кг корма), что, очевидно, связано с частыми голодовками зверей.
Печень белого медведя содержит необычайно много витамина А (до 9 - 10 тыс. мг/г) - во много раз больше, чем печень других животных. Для жира, находящегося в печени белого медведя, характерно очень высокое содержание жирных кислот, но относительно низкое содержание фосфолипидов и холестерина.
Молоко у самок очень густое, имеет запах рыбьего жира. Оно содержит (в %) 44,1 сухого вещества, 1,17 золы, 31 жира, 13 сухого вещества (без жира), 0,49 лактозы и 10,2 протеина. По химическому составу и чрезвычайно высокой жирности оно приближается к молоку китообразных и ластоногих.
У белого медведя уровень метаболизма выше, чем у бурого. Обнаружена необычайная стойкость к воздействию низких температур не только за счет его совершенной терморегуляции, по и благодаря низкой критической температуре. Даже при -50В° у белого медведя не происходит заметного повышения уровня газообмена, т.е. невозникает необходимости в использовании физиологического (ВлхимическогоВ») механизма терморегуляции, связанного с большим расходованием энергии.
Для головного мозга белого медведя характерно более умеренное, чем у бурого, развитие передних долей больших полушарий, большее развитие зрительной области и наличие менее объемистых обонятельных долей. Это позволяет предположить наличие у белого медведя более совершенного зрения, но более слабого обоняния, нежели у бурого медведя. Вообще головной мозг этого животного заметно отличается от мозга других хищных как своими очертаниями, так и более сложной топографией борозд и извилин больших полушарий. В этом отношении головной мозг белого медведя сходен с мозгом представителей отряда ластоногих, например гренландского тюленя и морского котика.
Глаза белого медведя небольшие, светло- или желто-коричневые (за счет коричневой радужины). Величина бинокулярного зрения у этого животного достаточно большая - около 180В°. Горизонтально-овальная форма роговицы и зрачка зверя увеличивает монокулярное поле его зрения. Угловая протяженность роговицы равна 105В° (у бурого медведя - 110В°), что свидетельствует о приспособлении глаз белого медведя к круглосуточной активности. Сетчатка белого медведя не обладает высокой светочувствительностью. Это тоже свидетельствует о сочетании признаков, характерных как для ночного, так и для дневного типа зрения. Белый медведь обладает относительно высокой способностью к восприятию объектов под водой.
3. АРЕАЛ и ЧИСЛЕННОСТЬ
Ареал белого медведя представляет кругополярную область, ограниченную северным побережьем материков, южным пределом распространения плавучих льдов и северной границей теплых морских течений. В пределах ареала можно выделить четыре области: постоянного современного обитания животных (более или менее peгулярного их появления, особенно в зимние месяцы); высокой численности; регулярного залегания в берлоги беременных самок; дальних заходов (или заносов со льдами) на юг (рис. 1).
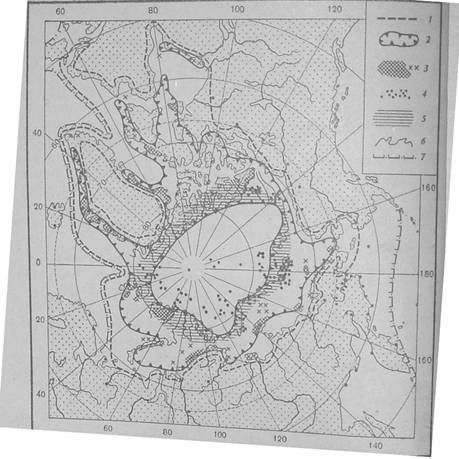
Рис. 1 - Распространение белого медведя в Арктике: 1 -южная граница области регулярных встреч; 2 - граница области высокой численности; 3- места залегания беременных самок в берлоги; 4 - места далеких заходов к югу (на суше) и встреч во льдах Северного Ледовитого океана; 5 - арктическое кольцо жизни; 6 - схематическое положение границы материкового склона; 7 - южная граница дрейфующих льдов зимой
I. Область постоянного современного обитания животных не выходит за пределы зимней границы дрейфующих льдов в северном полушарии. Она простирается до 50В° с.ш. на северо-востоке Канады и до 79В° с.ш. в Гренландском море, у западного побережья Шпицбергена.
Белые медведи распространены на всем побережье Гренландии, включая ее южную оконечность. Во льдах Гренландского моря они обитают к югу до о-ва Ян-Майен, в Баренцевом море - на островах Шпицбергена, Земли Франца-Иосифа, Новой Земли, на о-вах Медвежий, Вай-гач и Колгуев (за исключением юго-западной части) и на массивах льдов, располагающихся между этими участками суши. В Карском море белые медведи встречаются повсеместно, кроме южных частей Байдарацкой губы, Енисейского залива и Обской губы. В морях Лаптевых, Восточно-Сибирском, Чукотском и Бофорта южная граница постоянного обитания их совпадает с побережьем материка. В Беринговом море она не опускается далеко к югу от о-ва Св. Лаврентия. В Канаде эта область охватывает п-ов Мелвилл, ограничена побережьем Гудзонова залива, восточным побережьем п-ова Ньюфаундленд и севером одноименного острова. Белые медведи регулярно обитают в Баффиновом заливе и Девисовом проливе. К северу (в высоких широтах Арктики) звери всех половых и возрастных групп, кроме самок с новорожденными медвежатами, распространены вплоть до Северного полюса.
II. Область высокой численности белых медведей приурочена к материковому склону Северного Ледовитого океана и на севере, по-видимому, совпадает с сто границами, что обусловлено гидрологическими и гидробиологическими причинами.
Издавна известно существование в высоких широтах Арктики более или менее постоянных значительных участков открытой воды- полыней: Великой Сибирской, Восточно-Таймырской, Гренландской и др. Как установлено Я.Я. Гаккелем в 1957 г., участки открытой воды закономерно приурочены в Северном Ледовитом океане к районам материкового склона, а также, по-видимому, к районам крупных подводных хребтов, что связано с усилением приливно-отливных течений, наличием разных по динамике масс льда (неподвижный припай и дрейфующие льды), перемешиванием поверхностных холодных вод с нижележащими атлантическими. Перемешивание вод способствует формированию здесь слабоконтинентального климата и повышению биологической продуктивности вод. Замкнутое кольцо открытой воды, располагающееся в Северном Ледовитом океане, играет в жизни позвоночных животных, в том числе и белого медведя, громадную роль и с полным основанием может быть названо арктическим кольцом жизни. К нему, в частности, приурочено распространение на земном шаре своеобразного арктического дельфина - нарвала. Здесь зимует ряд видов арктических птиц, в их числе розовые чайки, некоторые чистиковые птицы; тут же наиболее многочисленны нерпы.
Существование арктического кольца жизни и тесная зависимость от него белого медведя подтверждается многочисленными наблюдениями советских дрейфующих станций ВлСеверный полюсВ». В тех случаях, когда станции находились в зоне этого кольца, полярники чаще встречали медведей. По мере того как станции удалялись к северу, встречи с медведями становились более редкими.
На юге область высокой численности белых медведей (в нее не входят участки морей с сильно разреженными льдами) ограничена не столь четко, как на севере. В Гренландии звери более обычны на северо-западе, севере и востоке острова. Они становятся многочисленными на севере Гренландского моря, на севере и востоке Шпицбергена, в центральных частях Баренцева моря, на северо-востоке Новой Земли. В восточной части Карского моря и на западе моря Лаптевых, где разреженных льдов практически не бывает, эта граница приближается к побережью материка, т.е. к южному пределу области постоянного современного обитания животных.
В архипелаге Новосибирских островов белые медведи более многочисленны на севере о-вов Анжу и на о-вах Де-Лонга. На востоке Восточно-Сибирского моря, в морях Чукотском и Бофорта граница области высокой численности зверей вновь приближается к побережьям материков. В Канаде она опускается далеко к югу, захватывая север и крайний юго-запад Гудзонова залива, север Баффинова залива и восток Баффиновой Земли.
III. Районы регулярного залегания в берлоги беременных самок. В отличиеот остальных особей вида, беременные медведицы имеют устойчивую связь с сушей, где осенью залегают в берлоги. Районы залегания в берлоги самок весьма постоянны, что связано с гидрологическими, климатическими, возможно, также кормовыми условиями, Эти районы располагаются преимущественно в пределах области высокой численности животных. В Гренландии к ним относятся участки, как северо-запада, так и северо-востока острова. В Баренцевом море очаги залегания в берлоги медведиц находятся на юго-востоке Шпицбергена и на западе Земли Франца-Иосифа. Далее к востоку регулярно, хотя и в небольших количествах звери размножаются на северном острове Новой Земли (как на его западной части - между Архангельской губой и заливом Русская Гавань, так и на восточной - между мысами Крашенинникова и Миддендорфа), на мелких островах Карского моря, на Северной Земле, на северном и северо-восточном побережьях п-ова Таймыр. В Восточной Сибири одиночные берлоги медведиц встречаются в дельте Лены, на Медвежьих островах, на побережье и прилежащих островах (Айон, Колючин) Чукотского полуострова. Более регулярно они залегают на севере Новосибирских островов. Здесь же, на о-ве Врангеля, находится один из крупнейших в Арктике очаг размножения животных.
На Аляске зимние убежища самок немногочисленны, Звери изредка размножаются в дельте реки Колвилл, у устья реки Купарук и иногда на прибрежном припае. В Канаде к районам регулярного залегания медведиц в берлоги относятся юг о-ва Банкса, побережье п-ова Симпсона, северо-восточное побережье Баффиновой Земли, северо-восток о-ва Саутгемптом. Районы регулярного залегания в берлоги беременных самок, следовательно, имеют небольшую общую площадь и размещаются очень неравномерно. Роль их в воспроизводстве запасов вида оказывается к тому же весьма неравнозначной, о чем могут дать представление приведенные ниже сведения о возможной общей численности берлог в советской Арктике:
Земля Франца-Иосифа 100-150
Новая Земля 50
Мелкие острова Карского моря 15
Северная Земля 20
Побережье Таймырского полуострова с прилежащими 30
Островами
Новосибирские острова 50
Материковое побережье Якутии с прилежащими островами 20
Материковое побережье Чукотского полуострова с прилежащими островами 30
О-в Врангеля не менее 200
Интересное открытие сделали американские зоологи: они обнаружили берлоги беременных медведиц на паковых льдах моря Бофорта.
IV. Область дальних заходов белых медведей на юг не имеет определенных границ, но она наиболее обширна. В подавляющем; большинстве случаев в районы, расположенные вне области их постоянного обитания, звери попадают вместе с плавучими льдами. Именно таким образом время от времени (преимущественно ранней весной) они достигают Исландии, в прошлом попадали на северные побережья залива Св. Лаврентия и Скандинавии, п-ов Канин. В пределах Тихого океана льды нередко заносят белых медведей в Анадырьский залив, к берегам Коряцкой Земли, иногда Камчатки, в центральные районы Берингова моря, а в прошлом даже до Курильских островов, берегов о-ва Хоккайдо и, возможно, о-ва Хондо (Хонсю). Непосредственно со льдами или пересекая Камчатку звери изредка попадают в Охотское и даже Японское моря.
Оказавшись после таяния льдов на побережье и стремясь вернуться в привычную обстановку, белые медведи нередко совершают большие переходы по суше, двигаясь иногда прямо на север. Очевидно, при таких обстоятельствах их чаще всего встречают в глубине континентов в Евразии и Северной Америке. Особенно многочисленны случаи встреч зверей вдали от моря на Чукотском полуострове, где медведи переходят из Берингова моря в Чукотское (например, из Мечигменской губы или бухты Руддера в Колючинскуку губу), в долинах рек Анадыря Белой и Пенжины (переходы в Чукотское море из Анадырского залива или из Охотского моря). Белые медведи заходят в глубь суши, на расстояния в сотни километров от побережья и в других районах: например, в центральную часть Таймырского полуострова, в долины рек Печоры и Хатанги, на Аляску.
Границы областей постоянного современного обитания белых медведей и их высокой численности претерпевают существенные сезонные колебания, связанные с изменениями в ледовом режиме Арктики. Летом, когда кромка плавучих льдов в Гренландском, Баренцевом и Беринговом морях, Девисовом проливе, Баффиновом, и Гудзоновом заливах сдвигается к северу, на юге остальных арктических морей образуются пространства открытой воды, становятся менее сплоченными льды Центральной Арктики - границы области практически сливаются. Общая площадь ареала сокращается, а распределение особей внутри его за счет продвижения животных в высокие широты бывает наиболее равномерным. Зимой белые медведи в большинстве районов спускаются к югу, исчезают из Центральной Арктики и концентрируются преимущественно у южного предела дрейфующих льдов и на арктическом кольце жизни. Площадь ареала вида в это время года становится наибольшей, а распределение зверей наименее равномерным.
В областях, испытывающих резкие колебания климата и ВлледовитостиВ» - потепления и похолодания Арктики, особенно на севере Атлантики (хотя в той или иной мере это относится ко всему ареалу вида), происходят многолетние изменения, как в численности, так и в распространении животных. В Гренландии, где до 1865 г., судя по результатам охоты, белые медведи были обычны в северо-западных районах и относительно редки в юго-западных, после 1865 г. количество зверей возросло на юго-западе и сократилось на северо-западе. После 1930 г. они стали редки на западе Гренландии, но обычны на востоке.
В целом площадь и границы области постоянного современного обитания белых медведей за последние несколько столетий изменились мало. Одним из доказательств может служить факт совпадения в большинстве районов Арктики южных границ этой области и зимнего распространения дрейфующих льдов. Исключение составляют лишь юг Баренцева моря (крайний северо-восток Мурманского побережья, п-ов Кании, побережья Тиманской, Малоземельной и Большеземельной тундр), горлоБелого моря и север Берингова моря, где, несмотря на благоприятные ледовые условия, звери отсутствуют, хотя в прошлом встречались регулярно.
Однако области высокой численности животных, регулярного залегания медведиц в берлоги и дальних заходов белых медведей на юг за последние 30-40 лет значительно сократились. Еще в 1900-1910 гг. белые медведи были многочисленны на западном побережье северного острова Новой Земли, на юге о-вов Анжу, в значительном количестве залегали в берлоги на Медвежьих островах и т.д. Падение численности этого вида связано с ростом населения и развитием индустрии в Арктике.
Как уже было отмечено, звери предпочитают держаться среди плавучих, но не мелкобитых льдов, перемежаемых с участками открытой воды, т.е. у кромки льда, у полыней и разводьев. Здесь круглый год бывают наиболее многочисленны нерпы - основной корм белого медведя. В этих условиях медведи успешнее на них охотятся.
При наличии открытой воды белых медведей можно встретить на льдах любого возраста. Полей очень молодого, непрочного льда толщиной до 5-6 см, лишенного на поверхности укрытий, они обычно избегают. Летом гладкие молодые льды, несомненно, привлекают зверей обилием нерп, образующих здесь линные залежки. В молодых льдах нерпы, а следовательно и медведи, более многочисленны в осенние и зимние месяцы. Одно- и двухлетние льды толщиной от 70 до 250 см, как правило, покрытые грядами торосов и спешными заносами, особенно благоприятны для размножения нерп (устройство подснежных нор, рождение и укрывание в них молодняка). Как свидетельствуют наблюдения, среди массивов этих льдов белые медведи держатся и охотятся главным образом весной. Среди многолетних льдов, имеющих толщину свыше 3 м и сглаженную многократным летним таянием поверхность (они встречаются преимущественно в Центральной Арктике), пет участков открытой воды, и белые медведи здесь относительно редки.
Намечается определенная зависимость между распределением животных, возрастом льда и географическим положением ледяных полей. По наблюдениям с самолета, в советской Арктике всюду, за исключением Баренцева моря, белых медведей можно чаще встретить на молодых льдах весной (март - май). Однако в Баренцевом море они тяготеют к старым льдам, разомкнутым в отличие от льдов Центральной Арктики.
Выходя на сушу, животные держатся в основном у морских побережий или вдали от моря по речным долинам. Заходить на ледники они, как правило, избегают, хотя изредка их встречают даже на ледниковом куполе Гренландии.
В берлоги самки залегают в основном вблизи морских побережий. По нашим наблюдениям, на о-ве Врангеля в 1964 г. зимние убежища самок (около 55%) располагались не далее 8 км от берега моря.
Возможность устройства медведицами родильных берлог зависит от условий накопления и таяния снега, а следовательно от рельефа местности: высоты склонов, их крутизны и экспозиции, преобладающего направления ветров в зимнее время, определяющих характер снежного покрова. В какой-то мере возможность залегания самок в зимние убежища зависит и от особенностей геологического строения суши.
Вследствие большой площади ареала белого медведя и крайне неравномерного распределения животных, работы, связанные с учетом этих зверей, дороги, поэтому современная их численность определена ориентировочно.
По данным Р. Скотта, впервые попытавшегося подсчитать общее количество медведей в Арктике, в 1956- 1958 гг. один белый медведь в среднем встречался на площади 83 км2. Общая численность зверей, обитающих у Аляски, определена в 2,5 тыс. особей, а численность на земном шаре - около 19 тыс.
Один из путей определения общей численности белых медведей, изучения ее динамики - использование данных советских ледовых авиаразведок, систематически проводимых па больших площадях от запада Баренцева моря до Берингова пролива. Анализ весенних наблюдений авиаразведки за 1962, 1967 и 1968 гг. показал, что в советской Арктике в апреле - мае на площадь 700 км2 приходится один белый медведь. При пересчёте на общую площадь льдов, составляющую по данным- 1968 г. 6 395 222 км2, количество белых медведей - л советской Арктике равно 5,6-6,6 тыс., а в пределах всего ареала - 10,7-13,6 тыс. особей.
Другим путем определения общего количества белых медведей может быть учет берлог беременных медведиц, особенно в местах их массового размножения. Основываясь на общей численности медведиц в СССР (500- 600) и считая, что беременные самки составляют около 20% популяций, автор в 1960-1965 гг. определил количество белых медведей в 5-10 тыс.
Приближенно о численности этих животных можно судить на основе анализа данных по их добыче и сведений о динамике и величине годового прироста. В конце 60-х - начале 70-х гг. во всей Арктике ежегодно добывалось по 1-1,5 тыс. медведей. Если в эти годы популяция их не возрастала и находилась в устойчивом состоянии, годовой прирост был близок 10%, а общее количество зверей составляло около 10 тыс.
Этот вопрос можно решать и комплексно: с одновременным привлечением данных авиаучетов, подсчета берлог и т.д. Идя по такому пути, Харингтон ориентировочно определил современную численность белых медведей, обитающих в Канаде, в 6-7 тыс., а во всей Арктике в 10 тыс. особей; по Ларсену общая численность белых медведей в Арктике составляет около 20 тыс. особей.
Динамика численности белых медведей обусловлена двумя основными факторами - антропогенным и климатическим. Как и большинство представителей фауны в земного шара, белые медведи ощущают на себе косвенное влияние человеческой деятельности. Рост в Арктике населения, совершенствование транспортных средств, развитие на арктических островах и побережьях индустрии, появление поселков и даже городов несомненно способствовали сокращению численности вида, ибо звери вытеснялись при этом с мест своего размножения. Возможно, что при участившихся контактах с домашними животными среди медведей распространились некоторые заболевания (особенно инвазии).
Однако человек гораздо сильнее влияет на численность белых медведей в результате охоты и истребления животных. Промысел белых медведей получил развитие с XVI-XVII вв., и несомненно, что уже к этому времени относится начало сокращения их численности. В XIX в. объем промысла значительно возрос, и стали появляться данные об уменьшении численности животных, в частности в районе Баренцева моря. Наибольших масштабов промысел достиг в XX в., особенно к 30-х гг., когда во всей Арктике, по нашим подсчетам, ежегодная добыча белых медведей стала превышать 2 тыс. особей. Если учесть, что эта величина явно превышала годовой прирост зверей, в тот период общее количество их составляло около 20 тыс. особей.
Падение численности продолжалось и позже, вплоть до середины 50-х гг., когда она достигла минимального значения за всю историю существования вида (по-видимому, менее 10 тыс. особей). Этот процесс был приостановлен в результате запрещения охоты на медведей в 1956 г. в советской Арктике. В настоящее время численность животных находится в относительно стабильном состоянии (или увеличивается).
Климатические факторы - периодически происходящие потепления и похолодания климата, по-видимому, оказывают на численность белого медведя существенное влияние. Последний период потепления, происходивший в первой половине XX в. и особенно резко проявлявшийся в 1920-1940 гг. в приатлантической части Арктики, характеризовался повышением температуры воздуха и морских вод (преимущественно в зимние месяцы), учащением зимой оттепелей и гололедиц, сокращением площади льдов в море. В Гренландии он вызвал отступление к северу холодолюбивой рыбы сайки - основного корма нерп, падение численности этого тюленя и, как естественное следствие, неравномерное в пределах острова сокращение численности белого медведя. Возможно, что на жизнь белых медведей зимой неблагоприятно сказывались также оттепели и гололедицы.
Наблюдения, сделанные как в зоопарках, так и в природе, свидетельствуют о примерно одинаковом соотношении полов между новорожденными медвежатами. В то же время существует основанное на результатах охоты мнение о преобладании среди взрослых особей самцов. По данным Л.Н. Попова, среди 69 медведей, добытых за три года на востоке Таймыра, 58 (84%) оказались самцами и лишь 11 (16%) -самками.
Фактические данные, характеризующие возрастной состав популяций белых медведей, оказываются весьма противоречивыми, что обусловлено географическим положением района, ледовыми условиями, сезоном, методикой сбора данных, и вряд ли могут быть использованы длякаких-либо конкретных выводов.
В советской Арктике, по сводным наблюдениям полярных станций, из 561 медведя на долю взрослых одиночек приходилось 63%, на долю самок с медвежатами разного возраста - 16 и медвежат - 21%.
4. ПИТАНИЕ
Основной корм белых медведей составляют несколько видов ластоногих, преимущественно нерпа и морской заяц. Кольчатая нерпа, или акиба (Phoca hispida),- наиболее распространенный и многочисленный в Арктике тюлень. Ареал его на юг простирается до о-ва Ньюфаундленд, Исландии, о-вов Ян-Майен и Медвежий, до северных побережий и приустьевых частей рек Восточной Европы, Азии, Северной Америки, включает Белое, Берингово и Охотское моря. Нерпа обитает во всей Центральной Арктике, вплоть до окрестностей Северного полюса. С наибольшей плотностью она населяет районы материкового склона Северного Ледовитого океана. Следовательно, области постоянного обитания и высокой численности белого медведя и нерпы совпадают. Для зверобоев медведи служат подчас своеобразным индикатором наличия и обилия в том или ином районе нерп.
Нерпа питается преимущественно рыбой, и распределение ее практически не зависит от глубины моря. Взрослые особи достигают в длину 180-200 см и веса 50-70 кг. Эти животные ведут относительно оседлый и одиночный образ жизни, тесно связаны с морскими, в основном разреженными, но не мелкобитыми льдами, на которых спят, линяют и рождают детенышей. Зимой, если поблизости нет открытой воды, нерпы проделывают в молодом льду ВлпродухиВ» - отверстия для дыхания. В торошеных льдах, имеющих на поверхности снежные наносы, нерпы проделывают более широкие отверстия - ВллазкиВ», через которые не только дышат, но и вылезают на лед (в подснежные убежища) для отдыха, рождения и кормления молодых. Продухи, а особенно лазки и зимние лежки нерп снаружи бывают незаметны, и медведи обнаруживают в них добычу но запаху (может быть, и на слух).
В марте-апреле самка рождает крупного детеныша, достигающего в длину 60 см и покрытого нежной белой шерстью. Белек нерпы в течение трех-четырех недель, пока он питается материнским молоком и линяет, лежит в убежище (но, если его потревожить, он, также как и мать, уходит в воду). Летом, с мая по август, линяют взрослые нерпы. В это время они подолгу спят на льду, преимущественно на ровных полях молодых льдов, у разводьев, промоин или лазок, нередко по нескольку особей. Однако сон их очень чуток. Периодически поднимая голову, они осматриваются и при малейшей опасности скатываются в воду.
Морской заяц, или лахтак (Erignatlius barbahis), достигает 260 см в длину и 200-300 кг веса. Он широко распространен в собственно арктических морях, но встречается реже, чем нерпа. Как и нерпа, морской заяц тесно связан со льдами, на которых отдыхает, линяет и размножается. В отлично от нее, обитает преимущественно на мелководьях (питается главным образом донными беспозвоночными) и, возможно, совершает сезонные миграции. Так же как и нерпы, морские зайцы в апреле - мае рождают детенышей в подснежных убежищах, устроенных среди торошеных льдов, в июле-августе линяют на ровных ледяных полях.
Морж (Odobaenus rosmarus) - крупнейшее из ластоногих животных Арктики - достигает в длину свыше 5 м и более 1,5 т веса. В пределах своего ареала - в арктических и субарктических морях - моржи образуют несколько самостоятельных стад, одно из которых круглый год обитает в морях Лаптевых и Восточно-Сибирском. Животные, принадлежащие к другим стадам, совершают сезонные миграции и на зиму откочевывают к кромке льдов - на север Тихого океана или Атлантики. Моржи питаются преимущественно донными беспозвоночными, поэтому приурочены главным образом к прибрежным или мелководным участкам морей. Тесно связаны они также со льдами (несплоченными), на которых образуют залежки. Во
Вместе с этим смотрят:
Анатомическое строение растений
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Анатомия человека
Анатомо-физологические механизмы безопасности и защиты человека от негативного воздействия
Бiологiчна роль кальцiю в органiзмi людини i тварин