Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Дыхание присуще всем живым организмам. Оно представляет собой окислительный распад органических веществ, синтезированных в процессе фотосинтеза, протекающих с потреблением кислорода и выделением диоксида углерода. А.С. Фаминцын рассматривал фотосинтез и дыхание как две последовательные фазы питания растений: фотосинтез готовит углеводы, дыхание перерабатывает их в структурную биомассу растения, образуя в процессе ступенчатого окисления реакционноспособные вещества и освобождая энергию, необходимую для их превращения и процессов жизнедеятельности в целом. Суммарное уравнение дыхания имеет вид:
C![]() H
H![]() O
O![]() Ва+ 6O
Ва+ 6O![]() Ва→ 6CO
Ва→ 6CO![]() Ва+ 6H
Ва+ 6H![]() O + 2875кДж.
O + 2875кДж.
Из этого уравнения становится ясно, почему именно скорость газообмена используют для оценки интенсивности дыхания. Оно было предложено в 1912 г. В. И. Палладиным, который считал, что дыхание состоит из двух фаз тАУ анаэробной и аэробной. На анаэробном этапе дыхания, идущем в отсутствие кислорода, глюкоза окисляется за счет отнятия водорода (дегидрирования), который, по мнению ученого, передается на дыхательный фермент. Последний при этом восстанавливается. На аэробном этапе происходит регенерация дыхательного фермента в окислительную форму. В. И. Палладин впервые показал, что окисление сахара идет за счет непосредственного окисления его кислородом воздуха, поскольку кислород не встречается с углеродом дыхательного субстрата, а связано с его дегидрированием.
Существенный вклад в изучение сути окислительных процессов и химизма процесса дыхания внесли как отечественные (И.П. Бородин, А.Н.Бах, С.П. Костычев, В.И. Палладин), так и зарубежные (А.Л. Лавуазье, Г. Виланд, Г. Кребс) исследователи.
Жизнь любого организма неразрывно связана с непрерывным использованием свободной энергии, генерируемой при дыхании. Неудивительно, что изучению роли дыхания в жизни растения в последнее время отводят центральное место в физиологии растений.
1. Аэробное дыхание
Аэробное дыхание тАУ это окислительный процесс, в ходе которого расходуется кислород. При дыхании субстрат без остатка расщепляется до бедных энергией неорганических веществ с высоким выходом энергии. Важнейшими субстратами для дыхания служат углеводы. Кроме того, при дыхании могут расходоваться жиры и белки.
Аэробное дыхание включает два основных этапа:
- бескислородный, в процессе, которого происходит постепенное расщепление субстрата с высвобождением атомов водорода и связыванием с коферментами (переносчиками типа НАД и ФАД);
- кислородный, в ходе которого происходит дальнейшее отщепление атомов водорода от производных дыхательного субстрата и постепенное окисление атомов водорода в результате переноса их электронов на кислород.
На первом этапе вначале высокомолекулярные органические вещества (полисахариды, липиды, белки, нуклеиновые кислоты и др.) под действием ферментов расщепляются на более простые соединения (глюкозу, высшие карбоновые кислоты, глицерол, аминокислоты, нуклеотиды и т.п.) Этот процесс происходит в цитоплазме клеток и сопровождается выделением небольшого количества энергии, которая рассеивается в виде тепла. Далее происходит ферментативное расщепление простых органических соединений.
Примером такого процесса является гликолиз тАУ многоступенчатое бескислородное расщепление глюкозы. В реакциях гликолиза шестиуглеродная молекула глюкозы (С![]() ) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С
) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С![]() ). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД
). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД![]() Ва(никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н
Ва(никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н![]() . НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты тАУ одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
. НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты тАУ одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
Н → Н![]() Ва+ е
Ва+ е![]() ,
,
а второй присоединяется к НАД или НАДФ целиком:
НАД![]() + Н + [Н
+ Н + [Н![]() + е
+ е![]() ] → НАД ∙ Н + Н
] → НАД ∙ Н + Н![]() .
.
Свободный протон позднее используется для обратного окисления кофермента. Суммарно реакция гликолиза имеет вид
C![]() H
H![]() O
O![]() Ва+2АДФ + 2Н
Ва+2АДФ + 2Н![]() РО
РО![]() Ва+ 2 НАД
Ва+ 2 НАД![]() →
→
2С![]() Н
Н![]() О
О![]() Ва+ 2АТФ + 2 НАД ∙ Н + Н
Ва+ 2АТФ + 2 НАД ∙ Н + Н![]() + 2 H
+ 2 H![]() O
O
Продукт гликолиза тАУ пировиноградная кислота (С![]() Н
Н![]() О
О![]() ) тАУ заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до CO
) тАУ заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до CO![]() и H
и H![]() O. Этот процесс можно разделить на три основные стадии:
O. Этот процесс можно разделить на три основные стадии:
1) окислительное декарбоксилирование пировиноградной кислоты;
2) цикл трикарбоновых кислот (цикл Кребса);
3) заключительная стадия окисления тАУ электронтранспортная цепь.
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А, в результате чего образуется ацетилкофермент а с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула CO![]() (первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н
(первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н![]() .
.
Вторая стадия тАУ цикл Кребса (рис. 1)
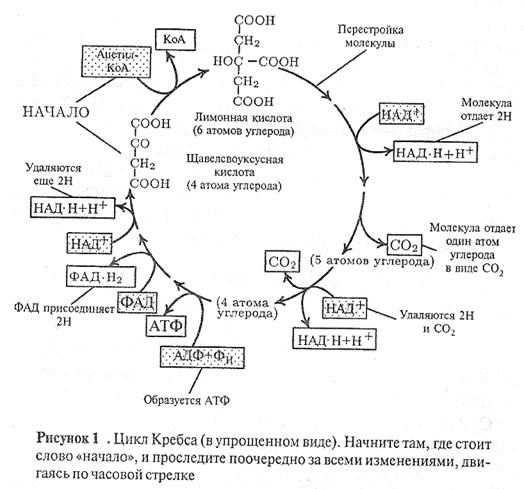
В цикл Кребса вступает ацетилтАУКоА, образованный на предыдущей стадии. АцетилтАУКоА взаимодействует со щавелево-уксусной кислотой, в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетилтАУКоА. В конце цикла щавелево-лимонная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетилтАУКоА, и цикл повторяется. Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + 3H![]() O + 3НАД
O + 3НАД![]() + ФАД + АДФ + Н
+ ФАД + АДФ + Н![]() РО
РО![]() →
→
КоА + 2CO![]() + 3НАД ∙ Н + Н
+ 3НАД ∙ Н + Н![]() +ФАД ∙ H
+ФАД ∙ H![]() + АТФ.
+ АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется 3CO![]() , 4 НАД ∙ Н + Н
, 4 НАД ∙ Н + Н![]() , ФАД ∙ H
, ФАД ∙ H![]() . Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
. Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
C![]() H
H![]() O
O![]() Ва+ 6 H
Ва+ 6 H![]() O + 10 НАД + 2ФАД →
O + 10 НАД + 2ФАД →
6CO![]() + 4АТФ + 10 НАД ∙ Н + Н
+ 4АТФ + 10 НАД ∙ Н + Н![]() + 2ФАД ∙ H
+ 2ФАД ∙ H![]() .
.
Третья стадия тАУ электротранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов, окисляются молекулярным кислородом до H![]() O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H
O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H![]() и ФАД ∙ H
и ФАД ∙ H![]() , передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н
, передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н![]() Ва+ 2е
Ва+ 2е![]() . Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
. Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
С помощью переносчиков ионы водорода Н![]() переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
|