Биосинтез антител. Структура и специфичность антигенов
Контрольная работа по биологии
2009
Содержание
1. Биосинтез антител
2. Структура и специфичность антигенов
3. Общая структурная характеристика молекул иммуноглобулинов
1. Биосинтез антител
Иммунная система, ответственная за биосинтез антител, состоит из ряда органов, основными из которых являются тимус, селезенка и периферические лимфоидные структуры в которых формируются три основных типа клеток: Т- и В-лимфоциты и макрофаги.
Антитела вырабатываются В-лимфоцитами, на поверхности которых уже имеются рецепторы, специфически связывающие антиген. В этот же комплекс включаются Т-лимфоциты и макрофаги. В результате межклеточной кооперации происходит активация В-лимфоцитов и их трансформация в плазматические клетки. Большая часть образовавшихся плазматических клеток синтезирует антитела, аналогичные по специфичности рецепторам на поверхности В-лимфоцитов, и секретирует их в кровь. Другая часть превращается в клетки Влиммунологической памятиВ», способные выделять антитела при повторном введении антигена.
Каждый В-лимфоцит содержит на поверхности около 100 тыс. рецепторов одинаковой специфичности. Антиген, встречаясь в кровотоке с комплементарным рецептором, проводит отбор соответствующего В-лимфоцита, который затем, трансформируясь в плазматическую клетку и многократно делясь, образует клон клеток. Эта теория биосинтеза антител, впервые сформулированная П. Эрлихом, а затем модифицированная в соответствии с уровнем развития науки Ф. Бернетом, получила название клонально-селекционной. Важно отметить, что каждый клон плазматических клеток секретирует гомогенные по своей структуре антитела. Однако так как антиген активирует в крови сразу несколько типов В-лрмфоцитов, которые содержат рецепторы различной степени специфичности по отношению к исходному антигену, такой иммунный ответ называется поликлональным, а антитела тАФ поликлональными.
Сыворотку животного, содержащую специфические к данному антигену антитела, называют антисывороткой. При этом обычно указывают, против какого антигена она выработана. Например, когда говорят об антисыворотке кролика против эритроцитов человека, подразумевают, что в ответ на введение в кровь кролика эритроцитов человека образуются специфические к ним антитела. Принципиально важным является то, что поликлональные антитела даже против одной-единственной антигенной детерминанты гетерогенны как по структуре активного центра, так и по физико-химическим свойствам. В том случае, если антиген поливалентен, например белок, то в сыворотке крови образуются антитела, направленные против каждой индивидуальной детерминанты, что еще более усложняет состав антител. Состав антител зависит от вида животного, а также стадии иммунного процесса.
Все перечисленные выше факторы влияют на гетерогенность антител и обусловливают определенные трудности как в изучении их структуры, так и в получении воспроизводимых стандартных препаратов антисывороток. Работы Келера и Мильштейна по гибридизации животных клеток открыли принципиально новый путь получения антител. Сущность метода заключается в том, что из организма иммунизированного животного выделяются лимфоциты, которые специальным образом ВлсливаютсяВ» с миеломными клетками. Образующиеся клетки получили название гибридом.
Особенностью таких клеток является их способность размножаться и продуцировать антитела в искусственных условиях вне организма. С помощью специальных методов клонирования можно выделить одну гибридную клетку, которая, размножаясь, будет секретировать в неограниченных количествах антитела только одного вида тАФ моноклональные антитела. Подчеркнем, что моноклональные антитела гомогенны как по специфичности, так и по физико-химическим свойствам. Вопросы, связанные с получением поликлональных и моноклоиальных а
2. Структура и специфичность антигенов
Понятие антигенная детерминанта включает в себя последовательность образующих ее химических функциональных групп и их пространственное расположение.
Белки. В молекулах белков антигенная детерминанта образуется совокупностью аминокислотных остатков. Размер антигенной детерминанты белков может варьировать от 5тАФ7 до 20 аминокислотных остатков. В случае длинных антигенных детерминант, по-видимому, только часть из них включается в собственно антигенную структуру, а остальные ответственны за ее конформацию.
Антигенные детерминанты белков бывают двух типов тАФ секвенциальные, т.е. представляющие из себя последовательность аминокислотных остатков в полипептидной цепи, и конформационные, образованные аминокислотными остатками из различных частей белковой цепи, но сближенные в пространственной конфигурации белковой глобулы. Оба типа антигенных детерминант-имеют важное значение для характеристики Влиммунного.портретаВ» белков. Во многих случаях единичная замена аминокислоты в структуре антигенной детерминанты или изменение конформации белковой глобулы являются достаточными для изменения антигенной специфичности макромолекулы.
Термин Влспецифичность антигенаВ» используется в двух смыслах: во-первых, как способность избирательно реагировать со специфическими антителами, во-вторых, поскольку белковые антигены поливалентны, как набор определенных антигенных детерминант. Если два антигена имеют только часть одинаковых антигенных детерминант, их называют перекрестно реагирующими антигенами. Примерами таких антигенов являются гомогенные белки близкородственных видов животных.
Некоторые антигенные детерминанты у белков с четвертичной структурой могут быть образованы фрагментами полипептидных цепей различных субъединиц, как, например, в иммуноглобулинах и гемоглобине. Как уже указывалось выше, антигенная детерминанта белков образована группой аминокислотных остатков, среди которых можно выявить иммунодоминантную группу. Примером такой группы является С-концевой аргинин декапептида белка вируса табачной мозаики.
Наиболее характерно эффект иммунодоминантной группы выявляется при модификации отдельных аминокислотных остатков химическими соединениями, например динитрофенильной группой. Антитела, образующиеся в такой модифицированной антигенной детерминанте, способны реагировать не только с ней, но и с другими аминокислотными остатками, к которым будет ВлпришитаВ» динитрофенильная группа. Более того, если в раствор, содержащий модифицированный белок и антитела, добавить свободный динитрофенол, то он будет ингибировать образование комплекса антиген тАФ антитело. Это явление, которое впервые было описано и детально изучено К- Ландштейнером, послужило основой для получения антител против различных гаптенов.
Существенный вклад в понимание структуры антигенных детерминант внесли работы по изучению антигенности синтетических пептидов. Классическим примером являются работы по синтезу пептидной петли лизоцима, которая является самостоятельной антигенной детерминантой. Было показано, что эта петля, содержащая внутреннюю дисульфидную связь, взаимодействует с антителами, получаемыми как против лизоцима, так и самой петли, однако образование комплекса не происходит, если дисульфидная связь восстановлена. Это свидетельствует о том, что структура антигенной детерминанты и ее специфичность зависят как от аминокислотной последовательности, так и от конформации данного фрагмента. Общим для антигенных детерминант является то, что они, как правило, находятся на поверхности белковой глобулы и образованы жесткими а-спиральными участками.
В работах М. Села было показано, что гомополимеры, состоящие из одинаковых аминокислот, не обладают антигенной структурой, в то время как гетерополимеры содержат антигенные детерминанты, причем чем сложнее состав полипептида, тем больше различных антигенных детерминант он образует. Важными здесь являются два фактора: жесткость конформации полипептидной глобулы и ее аминокислотный состав.
Существует несколько путей исследования структуры антигенных детерминант. Один из них основан на искусственном синтезе фрагментов первичной структуры, в которых проводятся замещения отдельных аминокислот. Другой путь тАФ изучение влияния избирательной модификации отдельных аминокислотных остатков. Еще один способ тАФ сравнение между собой гомологичных антигенов. Эффективным методом является также использование ограниченного протеолта и выявление фрагментов, содержащих антигенные детерминанты. Хотя ни один из этих путей не дает однозначных результатов, однако совокупность данных позволяет построить Влантигенный портретВ» данного белка.
Исключительную важность знания такого ВлпортретаВ» для иммунохимического анализа можно продемонстрировать на примере изучения антигенных детерминант раково-эмбрионального антигена и антигенно родственных ему белков. РЭА представляет гликопротеид с Мг=180 000, который содержит по массе около 50% углеводов. Он имеет 9 различных эпитопов, большинство из которых конформационно зависимые пептидные детерминанты. В сыворотке крови содержится еще целая группа белков, перекрестно реагирующих с РЭА, что затрудняет использование этого антигена для диагностических целей. В настоящее время с помощью моноклональных антител выделены три антигенные детерминанты, которые являются абсолютно специфичными для РЭА.
Знание антигенной специфичности является исключительно важным условием для создания диагностических иммунохимиче-ских наборов, так как позволяет проводить определение данного вещества в присутствии близкородственных соединений.
В роли антигенов могут выступать и сами иммуноглобулины. В этом случае организм вырабатывает антитела, которые часто называют вторичными. Например, при иммунизации кролика иммуноглобулинами другого вида животного вырабатываются вторичные антитела. В иммуноферментном анализе существует большая группа методов, в которых используют меченные ферментом вторичные антитела.
Обобщая данные о структуре антигенной детерминанта белков, можно выделить ее следующую характерную особенность: жесткий участок поверхности белковой глобулы, образованный одним или несколькими фрагментами полипептидной цепи, содержащими иммунодоминантную группу.
Нуклеиновые кислоты. Структура антигенных детерминант нуклеиновых кислот остается до сих пор малопонятной. Это обусловлено тем, что сами по себе нуклеиновые кислоты практически не иммуногенны, но в комплексе с белками к ним могут быть полу-чены антитела. Как и в случае белков, важную роль играет жесткость структуры полинуклеотида. В состав антигенной детерминанты входят три- и тетрануклеотиды. Они могут быть образованы как двуспиральными, так и односпиральными участками. Понимание структуры антигенных детерминант имеет важное значение не только в плане иммунохимической диагностики различных патологических процессов, но и в связи с активным развитием гибридизации ДНК, в которых антитела выступают в роли детектирующих систем.
Полисахариды. Полисахариды тАФ весьма сложная по своему составу и строению группа антигенов, входящая в структуру стенок микроорганизмов и многих других клеток и определяющая их специфичность. Полисахариды входят в состав многих белков и также играют роль антигенных детерминант. В большинстве случаев полисахаридные антигены представляют собой длинную цепь, к которой присоединены боковые короткие олигосахариды, содержащие 4тАФб остатков Сахаров, фактически являющиеся антигенными детерминантами. В одних случаях олигосахаридные группировки идентичны и тогда весь полисахаридный комплекс представляет собой чередующиеся антигенные детерминанты. В других случаях боковые цепи могут сильно различаться между собой. Как правило, в антигенную детерминанту входят остатки Сахаров, расположенные на конце боковых цепей, содержащие заряженную группу, которая является иммунодоминантной группой.
На специфичность олигосахаридных антигенных детерминант в структуре белков оказывает влияние ближайшее окружение по-липептидиой цепи белка-носителя и тип связи между сахарами.
В случае полисахаридных гомополимеров, таких, как декстран, антигенная детерминанта может состоять из фрагментов, содержащих до восьми остатков.
Липиды, входящие в состав комплексов со многими полисахаридами, не обладают антигенной специфичностью, по-видимому, из-за высокой конформационной подвижности. Таким образом, структура полисахаридных антигенных детерминант представляет собой олигосахаридные цепи длиной 4тАФ6 остатков, специфичность которых определяется химическим составом, типом гликозидных связей и остатками, находящимися в ближайшем окружении.
Гаптены. Гаптены тАФ это вещества, которые сами не вызывают иммунного ответа, но, будучи конъюгированы с носителями, обладают способностью стимулировать синтез против них антитела. Обычно принято считать, что гаптены тАФ это низкомолекулярные соединения, однако это не совсем верно. Например, нуклеиновые кислоты, полипептиды D-аминокислот имеют высокую молекулярную массу, но антитела против иих возникают только после коНъюгирования их с белками. В этом случае гаптены в конъюгатах с белком выступают в роли иммунодоминантной группы и поэтому в дальнейшем могут взаимодействовать с антителами независимо от белка носителя.
Структура и антигенная специфичность гаптенов определяется целым рядом факторов. В качестве гаптенов могут выступать самые разнообразные органические вещества с Мг>100. С практической точки зрения важными являются стероидные и пептидные гормоны, широкий круг лекарственных соединений, пестициды, различные продукты промышленного органического синтеза, обладающие аллергенным действием.
Антигенная специфичность гаптенов сильно зависит от их химической структуры. Так, введение дополнительных групп может сильно исказить Влантигенный портретВ» того или иного соединения. Например, тироксин и трийодтиронин отличаются только одним остатком I, чего вполне достаточно, чтобы антитела против этих гормонов сильно различались перекрестной реактивностью. Классическими примерами стали исследования с пара-, орто- и мета-аминобензойной кислотами, которые практически не дают перекрестных реакций при сопоставимых концентрациях.
Важным моментом является стереоспецифичность гаптенов; антитела против олигопептидов из D-аминокислот не реагируют с олигопептидами из L-аминокислот.
На антигенную специфичность сильное влияние оказывают аминокислотный остаток белка носителя, к которому пришит гап-тен, а также молекулярные размеры гаптена. Так, в длинных оли-гопептидных гаптенах замена аминокислотных остатков, которые расположены близко к белку-носителю, оказывает меньшее влияние, чем в коротких. Напротив, замены в удаленных от носителя аминокислотных остатках оказываются драматическими для антигенной структуры независимо от размеров гаптена. Аналогичны з-акономерности для олигосахаридов.
Антигенная специфичность гаптенов зависит не только от их химической структуры, но и от способа пришивки к белку-носителю, в частности от того, какая функциональная группа гаптена была использована для конъюгирования. Часто, при получении конъюгатов для иммунизации гаптены пришивают не непосредственно к молекуле белка-носителя, а через пространственную ВлножкуВ», содержащую обычно 4тАФ6 углеродных атомов. В этом случае сама ВлножкаВ» в комплексе с гаптеном выступает в качестве составной антигенной детерминанты и образующиеся антитела могут обладать меньшей эффективностью связывания с нативным гаптеном, чем с таким связанным через ВлножкуВ» гаптеном.
В некоторых видах иммуноферментного анализа в качестве одного из реагентов используют гаптен, меченный ферментом. Если связывание в таком конъюгате аналогично способу пришивки в конъюгате гаптена с белком-иосителем для получения антител, то говорят, что в анализе используют гомологичные антитела. Если же структура ВлножкиВ» и способ пришивки гаптена в обоих случаях различны, то говорят о гетерологичных антителах. Применение того или иного вида антител или конъюгатов в иммуноферментном анализе может весьма сильно сказываться на его чувствительности и некоторых других характеристиках.
Весьма существенным фактором для специфичности является химическая структура ВлнодаскыВ», в частности ее длина и ближайшее окружение гаптена. Все эти моменты крайне важно учитывать при разработке методов иммунохимического анализа гаптенов.
Сильное влияние различных факторов на антигенную структуру и специфичность гаптенов, по-видимому, объясняется их ограниченными размерами и особенностями структуры активных центров антител.
Знание антигенной структуры и специфичности гаптенов имеет важное значение для создания методов иммунохимического определения различных физиологически активных соединений, так как многие из них претерпевают различные биохимические превращения, в результате чего образуется группа близкородственных метаболитов.
3. Общая структурная характеристика молекул иммуноглобулинов
Антитела в организме выполняют две основные функции. Первая тАФ это распознавание и специфическое связывание соответствующих антигенов, вторая тАФ эффекторная, заключающаяся в индукции важнейших физиологических процессов, направленных на уничтожение антигена: лизис чужеродных клеток через активацию системы комплемента, стимуляция специализированных иммунокомпетентных клеток, выделение фармакологически активных веществ и т.д.
Развитие иммунохимии в течение последних 25 лет позволило установить строение антител, выявить стереохимические основы их функционирования. Особое внимание было уделено изучению структуры активных центров антител, что привело к созданию полицентровой модели связывания антигена. Исследования динамических структурных свойств иммуноглобулинов способствовало установлению характера связи между антигенсвязывающими и эффекторными функциями.
Иммуноглобулины по своей химической структуре относятся к большому классу природных соединений тАФ гликопротеидам, высокомолекулярным соединениям, состоящим из последовательности L-аминокислот, соединенных между собой пептидными связями.
Отдельные аминокислоты отличаются между собой боковыми заместителями R. В состав белков, в том числе иммуноглобулинов, входят двадцать аминокислотных остатков.
Полипептидная цепочка за счет образования водородных связей между карбонильным атомом кислорода и атомом водорода аминогрупп отдельных аминокислотных остатков способна определенным образом укладываться в пространстве, образуя так называемые а-спиральные участки и структуру. Такая локальная упорядоченная конформация отдельных участков полипептидной цепи получила название вторичной структуры.
В целом вся полипептидная цепь образует компактную трехмерную структуру тАУ третичную структуру: В одном растворе молекула белка сворачивается так, чтобы неполярные, или гидрофобные, боковые цепи аминокислотных остатков находились во внутренней, малодоступной для молекул воды области, а полярные, или ионизированные, группы образовывали внешний контактирующий с водой слой. Такое расположение аминокислотных остатков полипептидной цепи является термодинамическим наиболее выгодным состоянием, причем следует отметить, что это сворачивание пептидной цепи является высокоспецифичным и обусловлено первичной структурой молекулы.
Помимо рассмотренных причин сворачивания белков, обусловленных так называемыми гидрофобными взаимодействиями, определенный вклад в' стабилизацию трехмерной структуры вносят дисперсионные силы Лондона, возникающие в результате комплементарного распределения электронных облаков отдельных, рядом расположенных атомов. Энергия этого типа взаимодействий сильно зависит от расстояния между атомами и максимальна при так называемом ван-дер-ваальсовом расстоянии контакта, равном сумме ван-дер-ваальсовых радиусов взаимодействующих атомов.
Дополнительный вклад в поддержание трехмерной структуры молекул белков дают водородные и электростатические связи между боковыми группами аминокислот, а также ковалентные связи между отдельными частями полипептидной цепочки, например дисульфидные, или SтАФS-связи, возникающие между двумя остатками цистеина.
Некоторые молекулы белков состоят из несвязанных между собой ковалентно отдельных субъединиц. Такая пространственная организация получила название четвертичной структуры белков.
Несмотря на огромное разнообразие антител и их гетерогенность, все они обладают некоторыми общими структурными элементами, обеспечивающими выполнение их основных функций.
По своим антигенным, эффекторным свойствам и структурным особенностям иммуноглобулины подразделяются на пять основных классов: IgM, IgG, IgA, IgD и IgE.
Таблица 1 - Основные аминокислоты, входящие в состав белков
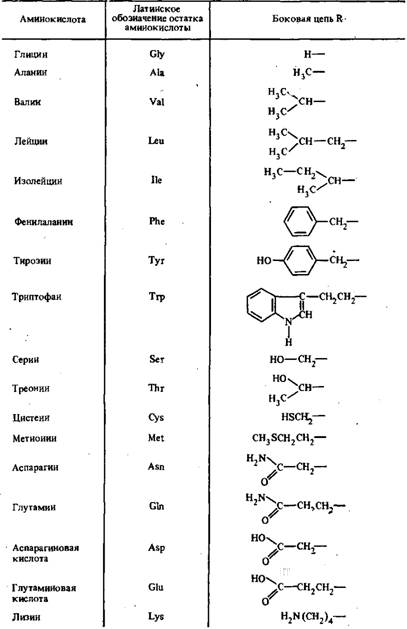
Общей структурной единицей всех иммуноглобулинов является комплекс из четырех полипептидных цепей тАФ двух идентичных между собой легких цепей с молекулярной массой 23000 каждая и тяжелый с молекулярной массой по 53000.
Каждая из лёгких цепей прочно соединена с концевыми участками тяжелых цепей благодаря наличию межцепочечных ди-сульфидных связей и множеству слабых гидрофобных, электростатических и других межатомных взаимодействий. Аналогичные связи существуют и между свободными участками тяжелых цепей. В целом структура такого комплекса напоминает латинскую букву ВлVВ» или ВлТВ» и характерна для иммуноглобулинов классов IgG, IgD и IgE.
При действии протеолитического фермента папаина молекула IgG распадается на три фрагмента, два из которых идентичны и сохраняют способность связывать антигены и третий, способный к кристаллизации. Именно Fc-фрагмент ответствен за эффекторную функцию антител тАФ связывание белка комплемента Clq, транспорт через мембраны, взаимодействие с мембранными рецепторами и т.д.
Другой протеолитический фермент пепсин "разрывает пептидную связь, расположенную ближе к СООН-концу цепи, от SтАФS-связи между Н-цепями в Fc-фрагменте. В результате образуются так называемый pFc-фрагмент, представляющий собой остатки тяжелых цепей, и соединенные дисульфидными связями два Fа-фрагмента, обозначаемые как ¥2-фрагмент. Последний также сохраняет способность к связыванию антигенов.
Антигенсвязывающий центр расположен в Н2-концевых частях Н- и L-цепей. Таким образом, каждая молекула IgG, а также Р2-фрагменты содержат по два антигенсвязывающих центра, а Fab-фрагмент тАФ один.
Необходимым условием для использования антител в иммуноферментном анализе является сохранение их способности специфически взаимодействовать с соответствующими антигенами.
Поэтому часто используют не целые молекулы антител, а только их фрагменты F2 или Fab, полностью сохраняющие эту способность. Такой подход позволяет в некоторых случаях устранить неспецифические реакции, обусловленные, в частности, взаимодействием Fc-фрагментов антител с поверхностями носителей.
Молекулы антител имеют большое число SтАФS-связей, которые можно разделить на 3 категории: межцепочечные связи, образуемые внутри структурной единицы между Н- и L-цепями и между Н- и Н-цепями, внутрицепочечные SтАФS-связи, возникающие в пределах одной и той же легкой или тяжелой цепи, и связи между Н-цепями отдельных четырехцепочечных комплексов, обусловливающих образование полимерных молекул тАФ IgM и IgA.
Структура иммуноглобулинов различных классов обусловлена числом и расположением SтАФS-связей в молекулах, а также количеством четырехцепочечных элементов. На рис. 2 приведено схематическое изображение молекул иммуноглобулинов различных классов.
IgM присутствует в сыворотке в виде пентамера четырехцепочечных комплексов, соединенных SтАФS-связями между Н-цепями. Некоторое количество IgA сыворотки также присутствует в виде димерной и тетрамерной формы. Полимерные IgA и IgM содержат Небольшую полипептидную цепь с Мг=14 000тАФ15000, которая, по-видимому, стабилизирует полимерную структуру. Кроме того, в димерный IgA входит секреторный компоненттАФбольшой гликопротеид с Мг=80 000.
Легкие цепи иммуноглобулинов бывают только двух типов тАФК или х. и являются общими для всех пяти классов, в то время как тяжелые цепи обладают структурными, иммунологическими и химическими особенностями, характерными для каждого класса иммуноглобулинов.
Для обозначения тяжелых цепей, относящихся к классам IgG, IgM, IgA, IgD и IgE, используют соответствующие греческие буквытАФ у, ц, а, б и е. На основании различий в химическом строении Н-цепей в пределах класса можно выделить подклассы иммуноглобулинов: 4 тАФ для IgG, 2 тАФ для IgA и 2 тАФ для IgM. В молекуле иммуноглобулина любого класса легкие цепи относятся только к одному типу.
Вместе с этим смотрят:
Анатомическое строение растений
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Анатомия человека
Анатомо-физологические механизмы безопасности и защиты человека от негативного воздействия
Бiологiчна роль кальцiю в органiзмi людини i тварин