Строение промежуточного мозга и его функции
I. Развитие и анатомическое строение промежуточного мозга
II. Функции промежуточного мозга
1. Таламус
2. Гипоталамус
3. Эпифиз
4. Ретикулярная формация ствола мозга
III. Заключение
Библиографический список
ВведениеОрганизм находится в непрерывном взаимодействии с окружающей его внешней средой. Это взаимодействие очень многогранно; оно обусловлено, с одной стороны, степенью сложности организации животного, а с другой - изменениями, которые постоянно происходят во внешней среде и в самом организме. Так как внешняя среда служит для организма не только источником, откуда он черпает материал для своего существования, но таит в себе различные для него опасности, то вполне понятно, что организм должен очень четко воспринимать различного рода раздражения и не менее четко на них реагировать. В связи с этим и развились высокодифференцированные органы нервной системы, приспособленные воспринимать и анализировать раздражения, идущие не только из внешней среды, но и из всех без исключения органов и тканей самого организма, и координировать деятельность организма в целом, проявляющуюся в его поведении, а также работу всех отдельных его органов и происходящий в них обмен веществ. Трофическая функция нервной системы впервые была выявлена И.П. Павловым. Такая координирующая функция осуществляется нервной системой при непременном участии органов чувств. Таким образом, интегрирующая функция принадлежит не только сосудистой системе, но в ещё большей степени нервной системе, влиянию которой подчинена и сама сосудистая система. Нервная система обеспечивает единство организма, взаимообусловленность всех его составных частей, единство организма и внешней среды, т.е. единство высшего порядка.
Основной структурной единицей нервной системы являются нейроны. Каждый нейрон состоит из тела и нервных отростков: рецепторного и эффекторного. Рецепторные отростки проводят раздражение к телу нейрона - это дендриты. Эффекторный отросток бывает только один; он проводит раздражение от тела нейрона к его периферии - это аксон ил нейрит.
В то время как нервные отростки служат только для передачи раздражений, тела нейронов выполняют необычно сложную функцию. В них воспринятое раздражение или затухает, если оно недостаточно сильно и действует монотонно, или же трансформируется и передаётся нейриту.
Весь процесс, протекающий в нервной клетке, от восприятия раздражения до ответа на него, т.е. до передачи раздражения с нервной клетки на выполняющий орган (мускульную или железистую клетку), называется рефлексом. В сложном организме рефлекс обычно осуществляется не одним нейроном, а рядом их, образующим цепь нейронов, или рефлекторную дугу.
I. Развитие и анатомическое строение промежуточного мозга
Промежуточный мозг - diencephalons - занимает довольно значительный участок головного мозга с обширной полостью третьего желудочка. Впоследствии, однако, полость желудочка становится щелевидной.
Пластинка покрышки служит сводом для третьего желудочка, который у всех животных остается зачаточным, состоящим из эпителиальной пластинки - lamina epithelialis, - которая, срастаясь с мягкой мозговой оболочкой, формирует сосудистую покрышку третьего мозгового желудочка - tela chorioidea ventriculi tertii, - заключающую в себе сосудистое сплетение. Покрышка внедряется отростками в полость третьего желудочка, а через межжелудочковое отверстие проникает также в конечный мозг, где и переходит в сосудистое сплетение боковых мозговых желудочков, - образовавшиеся за счет пластинки покрышки конечного мозга.
Производными свода являются:
1) непарный трубкообразный вырост - эпифиз и 2) парный - узел уздечки.
Эпифиз, или шишковидная железа, - epiphysis - рудимент третьего, так называемого теменного, глаза. Эпифиз, имеющийся почти у всех животных, развит не у всех одинаково и отсутствует лишь у немногих животных (у сумчатых и у некоторых других).
У млекопитающих эпифиз становится железой с внутренней секрецией. Он прикрепляется к зрительным буграм посредством двух ножек, на которых находятся ганглиозные утолщения - узел уздечки. Последние соединяются с обонятельными центрами, а также с ядрами тройничного нерва.
Боковые стенки третьего желудочка утолщаются в зрительные бугры - talami optici - вследствие вторичного образования ядер серого вещества и увеличения проводящих путей. Зрительные бугры играют роль важного промежуточного центра для проводящих путей, идущих в кору головного мозга и обратно. Оба бугра уже у рептилий соединяются друг с другом посредством промежуточной массы, состоящей из серого вещества; она проходит через полость третьего желудочка, вследствие чего последняя превращается в кольцевидный канал.
Производные базальной стенки мозгового пузыря, т.е. пластинки дна, объединяются под названием подталамическая часть - hipotalamus; она состоит из следующих органов.
Впереди зрительного перекреста вентральная стенка промежуточного мозга дает зрительный выступ - recessus opticus, - передняя стенка которого, переходящая в переднюю мозговую спайку, образована кольцевой пластинкой. Позади зрительного перекреста лежит другой непарный тонкостенный выступ в виде воронки - infundibulum. Передняя стенка её утолщается в серый бугор, - а сзади к ней примыкает сосцевидное тело - corpus mammilare, - также из серого вещества. В них оканчиваются волокна из свода в виде передних ножек последнего и из зрительных бугров.
Гипофиз, придаток мозга, - hypophysis - прилежит вентрально к воронке; он состоит из трех частей неодинакового происхождения, строения и различных по функции. Из эктодермы глотки первоначально образуется кармашкообразное выпячивание (карман Ратке), которое затем обособляется от стеки глотки и в виде пузырька примыкает к области воронки. Эпителий стенок пузырька образует разветвлённую железу. Затем просвет железы исчезает, но остаются тяжи от железистых клеток, окружённые большим количеством кровеносных сосудов. Ещё позднее обособляется промежуточная доля гипофиза, непосредственно граничащая с полостью воронки. У наземных животных за счет стенки воронки возникает нервная часть гипофиза, состоящая из нервных клеток. Таким образом, гипофиз у высших животных состоит из трех частей: дорзальной - нервной - neirohypohpysis, - вентральной - железистой - adenohypohpysis - и промежуточной. Железистая часть выделяет инкрет непосредственно в кровеносные сосуды (в кровь), а промежуточная и нервная - в третий мозговой желудочек.
У низших позвоночных животных - анамний - промежуточный мозг не играет еще такой роли, как у амниот, поэтому он и развит у них относительно слабо. Лишь с перемещением в него нервных центов из среднего мозга, обусловленным переходом к наземному образу жизни, промежуточный мозг начинает увеличиваться, оставляя далеко за собой средний мозг, что особенно заметно у человека. Благодаря наличию значительного количества ядер серого вещества промежуточный мозг становится центром корреляции для множества проводящих путей, идущих в кору головного мозга и обратно; отсюда понятно, что дифференциация промежуточного мозга начинается с момента роста конечного мозга.
II. Функции промежуточного мозгаПромежуточный мозг расположен между средним и конечным мозгом, вокруг третьего желудочка мозга. Он состоит из таламической области и гипоталамуса. Таламическая область включает в себя таламус, метаталамус и эпиталамус (эпифиз). Многие физиологи метаталамус объединяют с таламусом.
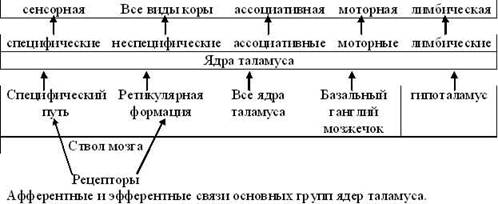
1. ТаламусТаламус (thalamus - зрительный бугор) представляет собой парный ядерный комплекс, составляющий основную массу (~20 г) промежуточного мозга и наиболее развитый у человека. В таламусе обычно выделяют до 60 парных ядер, которые в функциональном плане можно разделить на следующие три группы: релейные, ассоциативные и неспецифические. Все ядра таламуса в разной степени обладают тремя общими функциями: переключающей, интегративной и модулирующей.
Релейные ядра таламуса (переключательные, специфические) делятся на сенсорные и несенсорные.
Сенсорные релейные ядра переключают потоки афферентной (чувствительной) импульсации в сенсорные зоны коры (рис.1). В них также происходит перекодирование и обработка информации.
Кора больших полушарий
Вентральные задние ядра (вентробазальный комплекс) является главным реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной петли и примыкающих к ней волокнам других афферентных путей, где переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии; при этом функционально более тонко организованные части тела (например, язык, лицо) имеют большую зону представительства. Импульсация из вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины (поля 1-3), в которой формируются соответствующие ощущения. Электростимуляция вентральных задних ядер вызывает парастезии (ложные ощущения) в разных частях тела, иногда нарушение "схемы тела" (искаженное восприятие частей тела). Стереотаксическое разрушение участков этих ядер используется для ликвидации тяжелых болевых синдромов, характеризующихся острой локализованной болью и фантомными болями.
Латеральное коленчатое тело способствует переключению зрительной импульсации в затылочную кору, где она используется для формирования зрительных ощущений. Кроме корковой проекции, часть зрительной импульсации направляется в верхние бугры четверохолмия. Эта информация используется для регуляции движения глаз и в зрительном ориентировочном рефлексе.
Медиальное коленчатое тело является реле для переключения слуховой импульсации в височную кору задней части сильвиевой борозды (извилины Гешля, поля 41,42).
Переключение в таламусе афферентной импульсации от вестибулярного аппарата, по мнению некоторых ученых, происходит в вентральном промежуточном ядре и проецируется в нижнюю часть постцентральной извилины (поле 3), по мнению других - в медиальном коленчатом теле с дальнейшей проекцией в кору верхней и средней височной извилин (поля 21 и 22).
В релейных ядрах обнаружено несколько типов нейронов. Таламокортикальные (релейные) нейроны, имеющие длинный аксон, непосредственно обеспечивают переключение поступающей афферентной импульсации на нейроны сенсорной коры. Особенностью этой передачи является её высокая точность с минимальным искажением входного сигнала. Регуляция передачи возбуждения через релейные ядра осуществляется с помощью тормозных и возбуждающих вставочных нейронов. (Тормозные интернейроны возбуждаются как от таламокортикальных нейронов ядра, осуществляя возвратное торможение, так и от корковых нейронов, проецирующихся в данное ядро - корковый контроль проведения возбуждения).
Несенсорные релейные ядра таламуса (передние и вентральные) переключают в кору несенсорную информацию, поступающую в таламус из разных отделов головного мозга.
Впередние ядра импульсация поступает в основном из сосочковых тел гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору, откуда аксонные связи идут гиппокампу и опять к гипоталамусу, в результате чего образуется нейронный круг, движение возбуждения по которому обеспечивает формирование эмоций ("эмоциональное кольцо Пейпеца"). В связи с этим передние ядра таламуса рассматриваются как часть лимбической системы и иногда обозначаются как "лимбические ядра таламуса".
Вентральные ядраучаствуют в регуляции движения, выполняя таким образом моторную функцию. В этих ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая после этого проецируется в моторную и премоторную кору. Через эти ядра таламуса происходит передача в моторную кору сложных двигательных программ, образованных в мозжечке и базальных ганглиях.
Наряду с корковыми проекциями релейных ядер каждое из них получает нисходящие корковые волокна из той же проекционной зоны, что создает структурную основу для взаиморегулирующих отношений между корой и таламусом.
Ассоциативные ядра таламуса принимают импульсацию не от проводниковых путей анализаторов, а от других ядер таламуса. Эфферентные выходы от этих ядер направляются, главным образом, в ассоциативные поля коры. В свою очередь, кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. Главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга. Электростимуляция ассоциативных ядер вызывает в коре мозга биоэлектрическую реакцию в виде вторичного ответа.
Подушка получает главную импульсацию от коленчатых тел и неспецифических ядер таламуса. Эфферентные пути от неё идут в височно-теменно-затылочные зоны коры, участвующие в гностических (узнавание предметов, явлений), речевых и зрительных функциях (интеграция слова со зрительным образом), а также в восприятии "схемы тела". Электростимуляция подушки доминантного полушария приводит к речевым нарушениям типа аномии (нарушение называния предметов); разрушение подушки вызывает нарушение "схемы тела", устраняет тяжелые боли.
Медиодорсальное ядро получает импульсацию от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола. Проекция этого ядра распространяется на ассоциативную лобную и лимбическую кору (поля 12, 18). Оно участвует в формировании эмоциональной и поведенческой двигательной активности, а также, возможно, в образовании памяти. Разрушение этих ядер устраняет у больных страх, тревогу, напряженность, страдание от боли, но приводит к возникновению лобного синдрома: снижение инициативы, безразличие, гипокинезия.
Латеральные ядра получают зрительную и слуховую импульсацию от коленчатых тел и соматосенсорную импульсацию от вентрального ядра. Интегрированная сенсорная информация от этих источников далее проецируется в ассоциативную теменную кору и используется в функции гнозиса, праксиса, формировании схемы тела.
Неспецифические ядра составляют эволюционно более древнюю часть таламуса, включающую парные ретикулярные ядра и интраламинарную (внутрипластинчатую) ядерную группу. Ретикулярные ядра содержат преимущественно мелкие, многоотростчатые нейроны "ретикулярного типа" и функционально рассматриваются как производное ретикулярной формации ствола мозга.
Неспецифические ядра имеют многочисленные входы от других ядер таламуса, а также и внеталамические: по латеральному спиноталамическому, спиноретикулоталамическому путям, текто - и тегментоталамическим путям, проводящим преимущественно болевую и температурную чувствительность. В неспецифические ядра поступает непосредственно или через ретикулярную формацию также часть импульсации по коллатералям от всех специфических сенсорных систем. Кроме того, в неспецифические ядра поступает импульсация из моторных центров ствола (красное ядро, черное вещество), ядер мозжечка (шатра, пробкообразного), от базальных ганглиев и гиппокампа, а также от коры мозга, особенно лобных долей. Неспецифические ядра имеют эфферентные выходы на другие таламические ядра, кору больших полушарий, а также нисходящие пути к другим структурам ствола мозга.
Благодаря этим связям неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единый функциональный комплекс. На кору мозга неспецифический таламус оказывает преимущественно модулирующее (изменяющее состояние) влияние. Разрушение неспецифических ядер не вызывает грубых расстройств эмоций, восприятия, сна и бодрствования, образования условных рефлексов, а нарушает только тонкое регулирование поведения. В связи с этим модулирующее влияние неспецифических ядер таламуса, обеспечивающее "плавную настройку" высшей нервной деятельности, считается их главной функцией.
2. ГипоталамусГипоталамус включает в себя преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные (мамиллярные) тела. Микроскопически в гипоталамусе выделяют от 15 до 48 парных ядер, которые подразделяются на 3-5 групп.
Большинство авторов выделяют в гипоталамусе три основные группы ядер:
1) передняя группа содержит медиальное преоптическое, супрахиазматическое, супраоптическое, паравентрикулярное и переднее гипоталамическое ядра;
2) средняя (или туберальная) группа включает в себя дорсомедиальное, вентромедиальное, аркуатное (дугообразное) и латеральное гипоталамические ядра;
3) задняя группа включает в себя супрамамиллярное, премамиллярное, мамиллярные ядра, задние гипоталамическое и перифорниатное ядра, субталамическое ядро Луиса.
Важной физиологической особенностью гипоталамуса является высокая проницаемость его сосудов для различных веществ, в том числе и для крупных полипептидов. Это обусловливает большую чувствительность гипоталамуса к сдвигам во внутренней среде организма и способность реагировать на колебания концентрации гуморальных факторов. В гипоталамусе по сравнению с другими структурами головного мозга имеется самая мощная сеть капилляров (1100-2600 капилляров/мм2) и самый большой уровень локального кровотока.
Ядра гипоталамуса образуют многочисленные связи друг с другом, а также с выше - и нижележащими структурами ЦНС. Главные афферентные пути в гипоталамус идут от лимбической системы, коры больших полушарий, базальных ганглиев и ретикулярной формации ствола. Основные эфферентные пути гипоталамуса идут в ствол мозга - его ретикулярную формацию, моторные и вегетативные центры, в вегетативные центры спинного мозга, от мамиллярных тел к передним ядрам таламуса и далее, в лимбическую систему, от супраоптического и параветрикулярного ядер к нейрогипофизу, от вентромедиального и аркуатного ядер к аденогипофизу, а также эфферентные выходы к лобной коре и полосатому телу.
Гипоталамус является многофункциональной системой, обладающей широкими регулирующими и интегрирующими влияниями. Однако важнейшие функции гипоталамуса трудно соотнести с его отдельными ядрами. Как правило, отдельно взятое ядро имеет несколько функций, а отдельно взятая функция локализуется в нескольких ядрах. В связи с этим физиология гипоталамуса рассматривается обычно в аспекте функциональной специфики его различных областей и зон.
Гипоталамус как высший центр интеграции вегетативных функций. Изучение физиологической роли гипоталамуса с начала XXв. и по настоящее время показало, что при раздражении или разрушении его структур, как правило, происходит изменение вегетативных функций организма. Многолетние исследования швейцарского физиолога В. Гесса (1928-1968) доказали наличие в гипоталамусе двух функционально различных зон регуляции вегетативной сферы.
Стимуляция задней области гипоталамуса вызывала комплекс вегетативных реакций, характерный для симпатической нервной системы: увеличение частоты и силы сердечных сокращений, подъём АД, повышение температуры тела, расширение зрачков, гипергликемию, торможение перистальтики кишечника и др. полученные данные свидетельствуют о роли заднего гипоталамуса в интеграции различных симпатических центров. Эта область была названа Гессом эрготропной системой мозга, обеспечивающей мобилизацию и расходование энергетических ресурсов организма при активной его деятельности.
Раздражение преоптической и передней областей гипоталамуса сопровождалось признаками активации парасимпатической нервной системы: урежением ритма сердца, снижением АД, сужением зрачков, увеличением перистальтики и секреции желудка, кишечника и др. Эта область гипоталамуса была обозначена Гессом как трофотропная система, обеспечивающая процессы отдыха, восстановления и накопления энергетических ресурсов организма. Однако в дальнейшем было показано, что эрготропная и трофотропная области перекрывают друг друга, и можно только говорить о преобладании их в заднем и переднем гипоталамусе соответственно. Таким образом, приведенные данные свидетельствуют об интеграции вегетативных (симпатических и парасимпатических) центров в пределах гипоталамуса.
Вместе с тем на уровне гипоталамуса происходит не только интеграция деятельности различных вегетативных центров, но и включения их как компонента в более сложные физиологические системы различных форм биологического поведения, направленного на выживание организма. Поддержания гомеостаза и сохранения вида. Например, вазомоторные вегетативные реакции, обеспечивающие саморегуляцию АД, осуществляются сосудисто-двигательным центром продолговатого мозга, тогда как вазомоторные реакции, связанные с терморегуляцией, эмоциями, агрессивно-оборонительным поведением, реализуются на уровне гипоталамуса и более высоких центров.
Гипоталамо-гипофизарная система. Кроме вегетативного канала регуляции внутренней среды организма гипоталамус имеет мощный гуморальный путь реализации своих эффекторов. В этом плане главную роль играет взаимосвязь гипоталамуса с гипофизом. Клетки многих ядер гипоталамуса обладают нейросекреторной функцией и могут превратить нервный импульс в эндокринный секреторный процесс. Можно выделить две главные эндокринные связи гипоталамуса с гипофизом: гипоталамо-аденогипофизарную и гипоталамо-нейрогипофизарную.
Гипоталамо-аденогипофизарная связь. В 70-х годах XX в. Было установлено, что гипоталамус осуществляет контроль над эндокринной функцией аденогипофиза с помощью пептидных гормонов, образуемых мелкоклеточными нейронами в ядрах передней (паравентрикулярное, супраоптическое, супрахиазматическое ядра) и средней (дорсомедиальное, вентромедиальное, аркуатное ядра) групп. В этих ядрах образуется два вида пептидов: одни стимулируют образование и выделение гормонов аденогипофиза и называются рилизинг-гормонами (или либеринами), другие тормозят образование гормонов аденогипофиза и называются статинами. Известны пять либеринов: гонадолиберин стимулирует секрецию лютеинизирующего и фолликулостимулирующего гормонов, кортиколиберин - секрецию АКТГ, тиролиберин - секрецию ТТГ и пролактина, соматолиберин - секрецию СТГ, меланолиберин - секрецию меланоцитостимулирующего гормона. Тормозят секрецию аденогипофизарных гормонов: соматостатин, пролактостатин, меланостатин.
Либерины и статины поступают путем аксонного транспорта в срединное возвышение гипоталамуса и выделяются в кровь первичной сети капилляров, образованной разветвлениями верхней гипофизарной артерии. Далее с током крови они поступают во вторичную сеть капилляров, расположенную в аденогипофизе, и действуют на его секреторные клетки. Через эту же капиллярную сеть гормоны аденогипофиза поступают в кровоток и достигают периферических эндокринных желез. Эта способность кровообращения гипоталамо-гипофизарной области обозначается как гипофизарная портальная система.
Гипоталамо-нейрогипофизарная связь. В начале 30-х годов XXв. было обнаружено, что крупноклеточные нейроны супраоптического и паравентрикулярного ядер гипоталамуса являются эндокринными нейронами, образующими два нонапептидных гормона: антидиуретический гормон (АДГ) и окситоцин. Эти гормоны посредством аксонного транспорта поступают и депонируются в окончаниях аксонов, образующих нейроваскулярные синапсы на капиллярах задней доли гипофиза. Потенциал действия эндокринного нейрона запускает механизм перехода гормонов в капиллярную кровь нейрогипофиза и далее в общий кровоток. Основными мишенями для действия АДГ являются дистальные канальцы и собирательные трубочки почек, в которых он увеличивает реабсорбцию воды (уменьшает диурез) и сокращение гладких миоцитов сосудов (происходит сужение сосудов). Поэтому АДГ называют также вазопрессином. Наряду с этим АДГ оказывает влияние на нейроны гипоталамуса, активируя центр жажды и питьевое поведение. Основным физиологическим эффектом окситоцина является усиление сократительной функции матки и гладкой мускулатуры протоков молочных желез (выделение молока).
Роль гипоталамуса в терморегуляции. Организм человека, как и высших позвоночных, способен поддерживать постоянную температуру внутренней среды ("ядра") тела, несмотря на изменения температуры внешней среды. Проведенные сто лет назад опыты с перерезкой ствола мозга показали, что главной структурой, ответственной за температурный гомеостаз, является гипоталамус. В гипоталамусе выделено два центра: теплоотдачи и теплопродукции.
Центр теплоотдачи локализован в передней и преоптической зонах гипоталамуса и включает в себя паравентрикулярные, супраоптические и медиальные преоптические ядра. Раздражение этих структур вызывает увеличение теплоотдачи в результате расширения сосудов кожи и повышения температуры её поверхности, увеличения отделения и испарения пота, появления тепловой одышки. Разрушение центра теплоотдачи приводит к неспособности организма выдерживать тепловую нагрузку.
Центр теплопродукции локализован в заднем гипоталамусе и включает в себя медиальные, латеральные и промежуточные мамиллярные ядра. Его раздражение вызывает повышение температуры тела в результате усиления окислительных процессов, тонуса мышц и появления мышечной дрожи, сужения сосудов кожи. Разрушение этих ядер приводит к потере способности поддерживать температуру тела при охлаждении организма.
Виды нейронов терморегуляторного центра гипоталамуса. С помощью точных электрофизиологических методик были выявлены нейроны, способные реагировать генерацией потенциалов действия на изменения температуры гипоталамуса - термочувствительные нейроны (термодетекторы). Пороговая величина изменения температуры для них может быть от 0,2 до 1,0 0С. Эти нейроны расположены в основном в центре теплоотдачи, где они составляют от 30 до 70 % нейронов. Значительно меньше их (около 10 %) в центре теплопродукции. Среди термодетекторов количество тепловых нейронов примерно в 3 раза больше, чем холодовых. Другим видом нейронов терморегуляторного центра являются интегрирующие нейроны, основная функция которых состоит в суммации температурных сигналов от различных частей организма: терморецепторов гипоталамуса, среднего, продолговатого и спинного мозга, кожи, внутренних органов. Эти нейроны также расположены преимущественно в центре теплоотдачи. Переработанная в них информация о тепловом состоянии организма поступает на следующий вид нейронов - эфферентные терморегуляторные нейроны, которые расположены в основном в заднем гипоталамусе. Одни из них регулируют интенсивность механизмов теплопродукции, другие - механизмов теплоотдачи. Еще одна группа нейронов - тормозные интернейроны осуществляет реципрокное отношения между эфферентными нейронами теплоотдачи и теплопродукции. В центре терморегуляции есть также нейроны, воспринимающие нетемпературные стимулы: влияние экзогенных (бактериальных, вирусных) и эндогенных (интерлейкин 1, простагландин Е1, фактор некроза опухолей) пирогенов.
Роль гипоталамуса в регуляции поведения. Наиболее сложным вариантом интегративной деятельности гипоталамуса является объединение отдельных жизненно важных функций в сложные комплексы, обеспечивающие различные формы биологически целесообразного поведения: пищевого, полового, питьевого, агресивно-оборонительного и др., направленные на выживание индивидуума и вида. В основе этого поведения лежит возникновение в организме биологических потребностей, которые приводят к формированию в гипоталамических (а также лимбических и корковых) структурах мотивационного возбуждения, что выражается эмоционально окрашенным стремлением человека к удовлетворению соответствующей потребности. Удовлетворение потребности происходит через поведение. Однако в осуществлении даже биологических форм поведения гипоталамус обеспечивает только базовые механизмы. Эмоциональный компонент поведения осуществляется с обязательным участием лимбической системы. Корковых структур, а социализция биологического поведения - с участием новой коры, особенно лобной доли.
Пищевое поведение. В практической медицине давно было показано, что патологические процессы (опухоль, кровоизлияния, воспаление) в области гипоталамуса вызывают резкие изменения пищевого поведения, т.е. увеличение потребления пищи, так и отказ от неё. Экспериментальные исследования Б. Анада и Дж. Дробека (1951) показали, что разрушение небольшой зоны в области латерального гипоталамуса приводило к отказу от приема пищи (афагия), воды (адипсия) и гибели животного от истощения. Электрическая стимуляция этой зоны активизировала пищевое поведение: поиск и прием избыточного количества пищи (гиперфагия), ожирение. Эта зона гипоталамуса была названа центром голода. Разрушение вентромедиальных ядер гипоталамуса приводило к таким эффектам, как стимуляция центра голода: поиск пищи, гиперфагия, ожирение. Электрическая стимуляция этих ядер, напротив, прекращала пищевое поведение и прием пищи даже у голодного животного. Эта часть гипоталамуса в зоне вентромедиальных ядер была названа "центром насыщения".
Часть нейронов пищевого центра обладает хеморецепторной чувствительностью к некоторым веществам (глюкозе, аминокислотам, жирным и органическим кислотам) и гормонам (инсулину, гастрину, адреналину и др.) крови, уровень которых влияет на их импульсную активность. Наиболее изучены глюкозочувсвительные нейроны гипоталамуса. В "центре голода" выявлены нейроны, которые тормозят свою импульсную активность при увеличении концентрации глюкозы в крови. В "центре насыщения" имеются нейроны, которые, напротив, усиливают свою активность в этих условиях.
Питьевое поведение. Электрическое раздражение точек гипоталамуса, расположенных сзади и наружно от супраоптических ядер (Б. Андерсон, 1958), вызывает резко выраженную активизацию питьевого поведения и потребления воды (полидипсию). Разрушение этой зоны гипоталамуса, названной "центром жажды", напротив, приводит к отказу от приема воды (адипсии), причем эти явления прямо не связаны с изменением секреции АДГ. В дальнейшем было показано, что часть нейронов "центра жажды" обладает осморецептивными свойствами и стимулируется повышением осмотического давления крови. На активность "центра жажды" влияют также импульсы от периферических (сосудистых и тканевых) осморецепторов и концентрация некоторых веществ крови, например АДГ.
Половое поведение. Клинические наблюдения и экспериментальные данные с разрушением или стимуляцией показали, что некоторые структуры гипоталамуса осуществляют регуляцию половой мотивации и поведения. Половая дифференцировка гипоталамуса по женскому или мужскому типу формируется в последние дни внутриутробного развития и первые дни после рождения, хотя реализуется она при половом созревании. В гипоталамусе мужского организма функционирует "тонический" половой центр расположенный в средней области (аркуатные и вентромедиальные ядра). Нейроны этих ядер, выделяя гонадолиберины, оказывают постоянное стимулирующее влияние на секрецию лбтеинизирующего и фолликулостимулирующего гормонов гипофиза. В свою очередь, концентрация половых гормонов крови сильно влияет на возбудимость секреторных центров гипоталамуса (повышение их концентрации тормозит секрецию гоандолиберинов и наоборот). В женском организме кроме "тонического" функционирует также "циклический" центр, расположенный в преоптической и передней областях гипоталамуса (особенно в супрахиазматических ядрах) и осуществляющий регуляцию менструального цикла.
В регуляции полового поведения участвуют зоны заднего гипоталамуса и медиальный пучок переднего мозга. Раздражение их в эксперименте с животными (Дж. Олдс и сотр., 1954) и при операциях у людей показало формирования чувств радости, удовольствия, сопровождающихся эротическими переживаниями. Эти структуры были названы "центром удовольствия". Возбудимость сексуальных центров гипоталамуса существенно зависит от корковых и лимбических влияний, рефлекторной стимуляции с эрогенных генитальных и экстрагенитальных зон, уровня половых гормонов.
Агрессивно-оборонительное поведение. Агрессивные и оборонительные реакции были получены при раздражении различных зон гипоталамуса: передней и задней, вентромедиальной и латеральной (В. Гесс, 1928). Вероятно, это связано с тем, что имеются различные виды агрессивности, направленные на борьбу за самосохранение, лидерство в группе, территорию и др. Типичный симптомокомплекс этих реакций: яркое проявление отрицательных эмоций (гнев, ярость, страх), резкие вегетативные эрготропные сдвиги, попытки к нападению или бегству. Вместе с тем агрессивное поведение может быть без эмоционального возбуждения ("холодная атака") или сопровождаться положительной эмоцией. Перерезка ствола мозга ниже гипоталамуса ликвидирует агрессивное поведение. При осуществлении агрессивно-оборонительных реакций гипоталамус взаимодействует с центральным серым веществом среднего мозга. В этой структуре были обнаружены "нейроны агрессии" (Д. Адамс, 1968), которые через гипоталамус запускают реакции агрессии (борьба, драка) и не возбуждаются при других реакциях. При повреждении этих нейронов резко увеличивается порог электрического раздражения гипоталамуса для вызова реакции ярости. На агрессивное поведение выраженное стимулирующее влияние оказывают андрогены, особенно тестостерон (пороговая величина 1-2 мкг/л).
Перерезка ствола мозга сразу выше гипоталамуса (У. Кеннон) показала, что у животных легко возникает агрессивное поведение с реакциями ярости и и вегетативными проявлениями симпатического характера. Однако они не направлены на конкретный объект ("ложная агрессия и ярость"). Эти данные свидетельствуют о важной роли вышележащих отделов мозга в формировании "осмысленного" агрессивного поведения. Большое значение в этом аспекте имеют некоторые структуры миндалевидного комплексависочной доли, например базолатеральные, раздражение которых резко активирует, и разрушение - тормозит агрессию. Ограничивающее действие на агрессивное поведение и соотношение его с морально-правовыми нормами оказывает лобная кора (в частности, орбитальные зоны).
Поведение "бодрствование - сон". Клиническое изучение больных с поражением гипоталамуса позволило сформулировать предположение (К. Экономо, 1918), что "центр сна" расположен в переднем гипоталамусе, а "центр бодрствования" - в заднем. Экспериментальные исследования с повреждением различных участков гипоталамуса (В. Гесс,. 1929-1954; С. Ренсон, 1979; Т.Н. Ониани, 1983) подтвердили мнение клиницистов. Электрическое разрушение преоптической области (тормозная система) гипоталамуса вызывало синхронизацию электроэнцефалограммы и поведенческий сон. В это время активирующая ретикулярная формация ствола реципрокно заторможена. Стимуляция заднего гипоталамуса, напротив, вызывала десинхронизацию ЭЭГ и пробуждение. Пробуждающий эффект заднего гипоталамуса увеличивается при возбуждении активирующей ретикулярной формации ствола мозга. Роль гипоталамуса не ограничивается только формированием механизмов сна и бодрствования. Выполняя роль внутренних часов, гипоталамус является водителем этого околосуточного ритма, а важнейшими структурами являются супрахиазматическое и, возможно, вентромедиальное ядра, разрушение которых нарушает периодичность многих циркадиальных ритмов. Регулируя околосуточные биоритмы, гипоталам
Вместе с этим смотрят:
РЖсторiя виникнення та розвитку масажу
Аборты
Аденовирусная инфекция
Азотные и кислородные ванны, нафталановая нефть
Акушерська операцiя - накладання акушерських щипцiв