Деятельность центральной нервной системы
1. Общий план строения центральной нервной системы
Нервная система тАУ это совокупность специальных структур, объединяющая и координирующая деятельность всех органов и систем организма в постоянном взаимодействии с внешней средой.
К центральной нервной системе (ЦНС) относятся головнойи спинной мозг, которые состоят из серого и белого вещества. Серое вещество спинного и головного мозга тАУ это скопление нервных клеток вместе с ближайшими разветвлениями их отростков. Белое вещество тАУ это нервные волокна, отростки нервных клеток, которые имеют миелиновую оболочку (она придает волокнам белый цвет).
Структурно-функциональной единицей нервной системы является нервная клетка тАУ нейрон. Формы и размеры нейронов разных отделов нервной системы могут варьировать, но для них характерно наличие тела и отростков тАУ одного длинного (аксона) и множества древовидных коротких (дендритов). Аксон проводит импульсы от тела нейрона к периферическим органам или к другим нервным клеткам. Функция дендритов тАУ проведение импульсов к телу нейронов от периферических рецепторов и других нейронов. Передача нервного импульса от одного нейрона к другому происходит в местах их контактов- синапсах.
нервный система нейрон мозг
2. Нейрон как основной структурный и функциональный элемент нервной системы
Нервная ткань состоит из нервных клеток тАУ нейронов и глиальных клеток (Шванновских клеток).
2.1 Строение, типы и развитие нейронов
Нейрон, или
нервная клетка - это функциональная единица
нервной системы
. Нейроны восприимчивы к раздражению, т.е. способны возбуждаться и передавать электрические импульсы от
рецепторов к
эффекторам
. По направлению передачи импульса различают:
1) афферентные нейроны (чувствительные, сенсорные нейроны
) - проводят информацию об ощущении (импульс) от поверхности тела и внутренних органов в мозг;
2) эфферентные нейроны (двигательные нейроны
) тАУ проводят импульс (ВлкомандыВ») от головного и спинного мозга ко всем рабочим органам;
3) вставочные нейроны (ассоциативные) тАУ анализируют информацию и вырабатывают решения.
Вставочный нейрон передает возбуждение от афферентного (чувствительного) нейрона на эфферентные, лежит в пределах ЦНС. Тела эфферентных нейронов находятся в ЦНС или на периферии тАУ в симпатических, парасимпатических узлах. Аксоны этих клеток продолжаются в виде нервных волокон к рабочим органам (произвольным тАУ скелетным и непроизвольным тАУ гладким мышцам, железам).
Каждый нейрон состоит из
сомы
(клетки диаметром от 3 до 100 мкм, содержащей ядро и другие клеточные органеллы, погруженные в цитоплазму) и отростков, обычно одного длинного отростка - аксона и нескольких коротких разветвленных отростков - дендритов
. По дендритам импульсы следуют к телу клетки, по аксону - от тела клетки к другим нейронам, мышцам или железам. Благодаря отросткам нейроны контактируют друг с другом и образуют нейронные сети и круги, по которым циркулируют нервные импульсы.
Рис.1
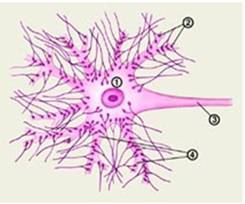
1 тАУ тело (сома) нейрона; 2 тАУ синаптические бляшки; 3 тАУ аксон нейрона; 4 тАУ дендриты.
На основании числа и расположения отростков нейроны делятся четыре группы:
1) униполярные нейроны
тАУ клетки с одним отрос
тком - аксоном. Истинных униполярных клеток в теле человека нет, если не считать униполярной формы нейробластов до периода образования дендритов.
2) биполярные нейроны
тАУ клетки с двумя отростками - аксоном и дендритом. Истинные биполярные клетки в теле человека встречаются редко. Такую форму имеют часть клеток сетчатки глаза, спирального ганглия внутреннего уха и некоторые другие;
3) псевдоуниполярные нейроны тАУ клетки, которые также имеют дендрит, идущий на периферию, заканчивающийся там чувствительными окончаниями (рецепторами), и аскон, несущий нервный импульс от тела клетки в ЦНС. Псевдоуниполярными они называются потому, что аксон и дендрит этих клеток начинаются от общего выроста тела, создающего впечатление одного отростка, с последующим Т-образным делением его.
4) мультиполярные (двигательные) нейроны. Из многих отростков такого нейрона один представлен аксоном, тогда как все остальные являются дендритами.
Нейрон развивается из небольшой клетки - предшественницы, которая перестает делиться еще до того, как выпустит свои отростки. Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется
конусом роста нервной клетки
. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков.
Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма ее может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении - некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединенными друг с другом мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных
актиновых филаментов
. Конус роста содержит также
митохондрии,
микротрубочки
и
нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счет добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости
медленного аксонного транспорта
в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста - это область быстрого
экзоцитоза
и
эндоцитоза, о чем свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком
быстрого аксонного транспорта
. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путем экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза
миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
2.2 Глиальные клетки
Помимо нейронов
нервная ткань
содержит клетки еще одного типа. Они выполняют опорную и защитную функции и называются
глиальными клетками или глией. По численности их в 10 раз больше, чем
нейронов
, и они занимают половину объема
ЦНС. Глиальные клетки окружают нервные клетки и играют вспомогательную роль. Глиальные клетки более многочисленные, чем нейроны: составляют по крайней мере половину объема ЦНС (
рис. 1-18
).
Глия не только выполняет опорные функции, но и обеспечивает многообразные метаболические процессы в нервной ткани, участвует в формировании миелиновой оболочки и способствует восстановлению нервной ткани после травм и инфекций.
Клетки нейроглии делятся на несколько типов.
Клетки эпендимы
выстилают
желудочки головного мозга и
спинномозговой канал
и образуют эпителиальный слой в
сосудистом сплетении. Они соединяют желудочки с нижележащими тканями.
Клетки макроглии
делятся на две категории -
астроциты и
олигодендроциты
.
Протоплазматические астроциты локализованы в
сером веществе
; от тела клетки, содержащей овальное ядро и большое количество
гликогена, отходят сильно разветвленные короткие и толстые отростки.
Фибриллярные астроциты
локализованы в
белом веществе. Ядро у них также овальное, и тело клетки содержит много гликогена
, но отростки длинные и менее разветвленные, некоторые ветви буквально упираются в стенки кровеносных сосудов. Эти клетки переносят питательные вещества из
крови в нейроны.
Астроциты двух типов взаимосвязаны и образуют обширное трехмерное пространство, в которое погружены нейроны. Они часто делятся, образуя в случае повреждений центральной нервной ситемы рубцовую ткань.
Олигодендроциты
локализованы в сером и белом веществе. Они мельче астроцитов и содержат одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек, а само оно содержит цитоплазму с большим количеством рибосом.
Шванновские клетки - это специализированные олигодендроциты, синтезирующие миелиновую оболочку
миелинизированных волокон
.
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. От каждого конца маленького продолговатого тела клетки, содержащей
лизосомы
и хорошо развитый
аппарат Гольджи, отходит по толстому отростку. От всех его ветвей отходят более мелкие боковые веточки. При повреждении мозга эти клетки превращаются в
фагоциты
и, перемещаясь при помощи амебоидного движения, противостоят вторжению чужеродных частиц.
2.3 Взаимодействие глии и нейронов
Между нейронами и глиальными клетками существуют сообщающиеся между собой щели размером 15-20 нм, так называемое
интерстициальное пространство
, занимающее 12-14% общего объема мозга.
Во время
потенциала действия концентрация ионов
калия
в интерстициальном пространстве может возрастать от 3-4 ммоль/л до 10 ммоль/л, что может вызвать значительную
деполяризацию нервных клеток. В результате же активного транспорта ионов калия его внеклеточная концентрация может стать ниже нормальной, что вызывает
гиперполяризацию нервных клеток
.
Глиальные клетки имеют высокую проницаемость для ионов калия. Когда несколько глиальных клеток деполяризуются вследствие местного повышения концентрации ионов калия, между деполяризованными и недеполяризованными клетками возникает ток, создающий вход ионам калия в деполяризованные глиальные клетки, в результате чего внеклеточная концентрация ионов калия уменьшается. Благодаря высокой проницаемости глиальных клеток для ионов калия и электрическим связям между ними глиальные клетки действуют как буфер в случае повышения внеклеточной концентрации калия. Данных об активном поглощении ионов калия путем ионного насоса в глиальных клетках нет, хотя, возможно, они активно поглощают нейромедиаторы в некоторых
синапсах, ограничивая таким образом время действия
медиатора
.
Рис.2
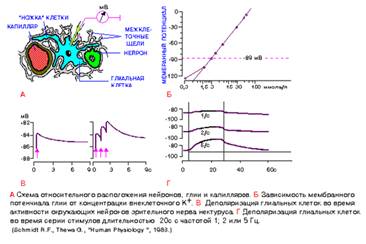
СВОЙСТВА ГЛИАЛЬНЫХ КЛЕТОК
А. Схема относительного расположения нейронов,
глии
и
капилляров, составленная по электронно - микроскопическим данным. Глиальная клетка - астроцит (на рисунке желтая), в которую введен микроэлектрод для регистрации
мембранного потенциала
, находится между капилляром и нейроном. Все клеточные элементы разделены межклеточными промежутками шириной порядка 15 нм (на схеме относительная ширина щелей увеличена).
Б. Зависимость мембранного потенциала глии (ордината) от концентрации внеклеточного К+ . Средний уровень
потенциала покоя
составляет -89 мВ. Экспериментальные данные соответсвуют величинам потенциала, рассчитанным по уравнению Нернста, за исключением данных в области концентраций калия ниже 0,3 ммоль/л.
В. Деполяризация глиальных клеток во время активности окружающих нейронов зрительного нерва. Стимулы (с интервалом 1 с) показаны вертикальными стрелками.
Г. Деполяризация глиальных клеток в том же препарате во время серии стимулов длительностью 20 с с частотой 1, 2 и 5 Гц, в последнем случае деполяризация достигала почти 20 мВ. Следует обратить внимание, что на рис. В и Г временной ход деполяризации гораздо медленнее по сравнению с потенциалом действия.
2.4 Синапс
Синапс (греч. synapsis соприкосновение, соединение) тАФ специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически синапс образован контактирующими мембранами двух клеток. Мембрана, принадлежащая отросткам нервных клеток, называется пресинаптической, мембрана клетки, к которой передается сигнал, тАФ постсинаптической.
Схема строения межнейронного синапса:
Рис.3

1 тАФ нервное волокно (аксон); 2 тАФ везикулы или синаптические пузырьки; 3 тАФ синаптическая щель; 4 тАФ рецепторы для медиатора; 5 тАФ постсинаптическая мембрана; 6 тАФпресинаптическая мембрана; 7 тАФ митохондрия.
В соответствии с принадлежностью постсинаптической мембраны синапсы подразделяют на нейросекреторные, нейромышечные и межнейроннные. Последние в зависимости от места их расположения разделяют на аксодендритические, аксосоматические, аксо-аксональные и дендро-дендритические. Различают
аксодендритные синапсы
(синапс между концевыми веточками
аксона одного нейрона
и дендритами другого нейрона) и
аксосоматические синапсы
(синапс между концевыми веточками аксона одного нейрона
и телом другого нейрона).
Рис.4
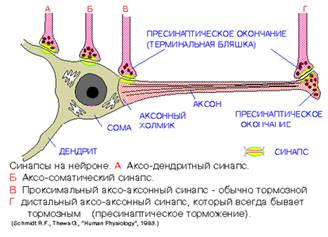
В зависимости от природы проходящих через синапсы сигналов, синапсы делятся на
электрические синапсы (так называемые эфапсы) и
химические синапсы
. В синапсах с химической передачей возбуждения между пре- и постсинаптической мембранами имеется синаптическая щель, куда выделяется химическое вещество-передатчик тАФ медиатор. Химические синапсы часто обозначают по названию медиатора (например, холинергические, адренергические, серотонинергические и т.п.). В эфапсе пре- и постсинаптические мембраны плотно соприкасаются и возбуждение передается посредством электрического тока. В зависимости от изменения биопотенциала постсинаптической мембраны различают синапсы деполяризующие, или возбуждающие, и гиперполяризующие, или тормозные.
Число синапсов очень велико, что обеспечивает большую площадь для передачи информации. На
дендритах и телах отдельных двигательных
нейронов спинного мозга
находится свыше 1 000 синапсов. Некоторые
клетки головного мозга могут иметь до 10 000 синапсов.
2.5 Механизм передачи возбуждения
Механизм передачи возбуждения принципиально одинаков во всех химических синапсах. В нем можно выделить следующие основные этапы: синтез и депонирование медиатора в пресинаптическом нейроне и его окончаниях; высвобождение медиатора из депонирующих везикул и его выход в синаптическую щель; взаимодействие медиатора со специфическими хеморецепторами постсинаптической мембраны с последующей генерацией биоэлектрического потенциала; инактивация выделенного медиатора с помощью ферментов или системы обратного поглощения. Биологически активные вещества, выполняющие функцию медиаторов, делят на несколько групп. К классическим нейромедиаторам относят ацетилхолин,адреналин и норадреналин, дофамин, серотонин, а также аминокислоты глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды. В соответствии с принципом Дейла, каждый отдельный нейрон в своих синаптических окончаниях один и тот же медиатор, поэтому нейроны можно обозначать также по виду медиатора: холинергические, адренергические, ГАМК-эргические, пептидергические. Синтез нейромедиаторов осуществляется как в соме нейрона с последующим аксонным транспортом, так и непосредственно в пресинаптических окончаниях аксона, где медиатор концентрируется в везикулах, или синаптических пузырьках.
В состоянии функционального покоя в пресинаптическом окончании происходит случайный контакт синаптических пузырьков с пресинаптической мембраной и выделение в синаптическую щель порции (кванта) медиатора из отдельной везикулы. Например, установлено, что в С. млекопитающих квант классического медиатора 10×ацетилхолина насчитывает 4210×тАФ44 молекул. Выделившийся в синаптическую щель медиатор взаимодействует с хеморецепторами постсинаптической мембраны и приводит к возникновению миниатюрного постсинаптического потенциала. Приходящий к пресинаптическому окончанию потенциал действия в несколько раз увеличивает количество выделяемого в синаптическую щель медиатора. Взаимосвязь между потенциалом действия пресинаптической мембраны и процессом выделения медиатора из везикул в синаптическую щель обеспечивается ионами Са++. Выделяющийся в синаптическую щель медиатор взаимодействует с различными хеморецептивными участками на постсинаптической мембране. Роль мембранных, или клеточных, рецепторов играют белковые молекулы, обладающие способностью ВлузнаватьВ» специфические для них вещества и вступать с ними в реакцию. В этом процессе большое значение придается системе аденилатциклаза-циклический АМФ. Предполагается, что взаимодействие медиатора с рецепторным участком аденилатциклазы приводит к образованию циклического АМФ с последующим повышением активности протеинкиназ цитоплазмы и ядра клетки. Вследствие этого интенсифицируется фосфорилирование белков, белковые молекулы подвергаются конформационным изменениям и происходит активация специальных ионных каналов мембраны. При увеличении проницаемости постсинаптической мембраны для ионов натрия, калия и хлора возникает ее деполяризация, регистрируется возбуждающий постсинаптический потенциал (ВПСП). При увеличении проницаемости лишь для ионов калия и хлора мембрана гиперполяризуется и регистрируется тормозный постсинаптический потенциал (ТПСП). ВПСП могут суммироваться, и при достижении величины деполяризации мембраны критического уровня генерируется потенциал действия. ТПСП тормозит генерацию потенциала действия, уменьшая суммарную величину ВПСП.
3. Строение и функции спинного мозга
Спинной мозг иннервирует скелетную мускулатуру (кроме мышц головы) и внутренние органы. Афферентными (чувствительными) путями спинной мозг связан с рецепторами, а эффернтными тАУ со скелетной мускулатурой и со всеми внутренними органами. Основные функции спинного мозга:
1.рефлекторная (принимает участие в двигательных реакциях)
- здесь располагаются центры безусловных рефлексов (коленный рефлекс);
- вегетативные центры рефлексов мочеиспускания, дефекации, рефлекторная деятельность желудка.
2.проводниковая (проведение нервных импульсов)
- осуществляется связь различных отделов спинного мозга;
- связь головного мозга с остальными частями ЦНС;
- соединение рецепторов с исполнительными органами.
По внешнему виду спиной мозг представляет собой продолговатый, несколько плоский цилиндрический тяж длиной 45 см, диаметром 1 см . Он расположен в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В центре спинного мозга находится канал, заполненный спинномозговой жидкостью. Спинной мозг состоит из 31-32 сегментов: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1-2 копчиковых. Каждому сегменту спинного мозга соответствует отдельный участок тела, который иннервируется от спинномозгового нерва определенного сегмента.
Рис.5
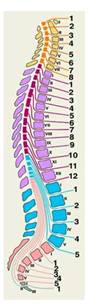
Схематическое изображение соотношений сегментов спинного мозга и позвонков на сагиттальном разрезе позвоночника. Оранжевым и желтым цветом обозначены шейные сегменты и шейные позвонки, фиолетовым и сиреневым тАФ грудные, голубым тАФ поясничные и копчиковые, розовым тАФ крестцовые. Римскими цифрами обозначены позвонки, арабскими тАФ корешки спинномозговых нервов соответствующих сегментов.
Вдоль всей передней поверхности спинного мозга в срединной сагиттальной плоскости тянется передняя срединная щель, а вдоль задней поверхности тАУ задняя срединная борозда, которые разделяют спинной мозг на две симметричные половины. На его поверхности находятся две передние латеральные борозды, из которых выходят передние корешки, а на задней поверхности есть задние латеральные борозды тАУ места входа с обеих сторон в спинной мозг задних корешков. Передние корешки заключают в себе только двигательные волокна, потому называются еще двигательными, задние - только чувствительные волокна, потому называются чувствительными корешками. Передние и задние корешки нервов проходят через межпозвоночные отверстия, где сливаются в один общий ствол, причем каждый задний корешок снабжается ганглиозным утолщением; нижние крестцовые пары выходят через передние и задние отверстия крестцовой кости. По выходе из позвоночника каждый нервный ствол разделяется на ветви, которые сперва имеют двигательные и чувствительные волокна и только при дальнейших разветвлениях дифференцируются, из 31-й пары нервов спинного мозга первые 8 пар называются шейными, следующие 12- грудными или спинными, 5 крестцовых и 1 копчиковая или хвостцовая. Между соседними нервами на некоторых местах образуются соединения посредством веточек, составляющие так наз. нервные сплетения. От этих сплетений и отходят в различные части тела главные нервные стволы, снабжающие эти части чувствительными или двигательными волокнами.
Серое вещество содержит нервные клетки и на поперечном разрезе напоминают форму бабочки. На протяжении всего спинного мозга серое вещество образует две вертикальные колонны, которые располагаются с двух сторон центрального канала. В каждой колонне различают передний и задний столбы (или рога). На уровне нижнего шейного, всех грудных и двух верхних поясничных сегментов спинного мозга в сером веществе выделяют боковой столб, который в других отделах спинного мозга отсутствует. Серое вещество задних рогов имеет неоднородную структуру. Основная масса нервных клеток заднего рога образует студенистое вещество и собственное ядро, а у основания заднего рога хорошо очерченное прослойкой белого вещества тАУ грудное ядро, которое состоит из крупных нервных клеток.
Рис.6
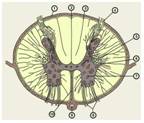
Поперечный разрез спинного мозга: 1 тАФ мягкая оболочка; 2 тАФ дорсолатеральная (задняя) борозда; 3 тАФ промежуточная дорсальная (задняя) борозда; 4 тАФ дорсальный (задний) корешок; 5 тАФ дорсальный (задний) рог; 6 тАФ боковой рог; 7 тАФ вентральный (передний) рог; 8 тАФ вентральный (передний) корешок; 9 тАФ передняя спинальная артерия; 10 тАФ вентральная (передняя) срединная щель.
Клетки всех ядер задних рогов серого вещества тАУ это, как правило, вставочные, промежуточные нейроны (интернейроны), отростки которых идут в белое вещество спинного мозга и далее к головному мозгу. Промежуточная зона, расположенная между передними и задними рогами, сбоку представлена боковым рогом. В последнем находятся центры симпатической части вегетативной нервной системы.
Белое вещество находится снаружи серого вещества. Борозды спинного мозга разделяют белое вещество на симметрично расположенные слева и справа три канатика: передний, боковой и задний. Белое вещество представлено отростками нервных клеток. Совокупность этих отростков в канатиках спинного мозга составляют три системы пучков тАУ трактов (проводников):
1) короткие пучки ассоциативных волокон, которые связывают сегменты спинного мозга, расположенные на разных уровнях;
2) восходящие (чувствительные, афферентные) пучки, направляющиеся к центрам головного мозга или к мозжечку;
3) нисходящие (двигательные, эфферентные) пучки, идущие от головного мозга к клеткам передних рогов спинного мозга.
В белом веществе задних канатиков находятся восходящие пути, а в передних и боковых канатиках проходят восходящие и нисходящие системы волокон.
Передний канатик включает следующие проводящие пути:
1) передний корково-спинномозговой (пирамидный) путь. Этот путь передает импульсы двигательных реакций от коры большого полушария головного мозга к передним рогам спинного мозга;
2) передний спинно-таламический путь тАУ обеспечивает проведение импульсов тактильной чувствительности;
3) преддверно-спинномозговой тАУ берет начало от вестибулярных ядер VIII пары черепных нервов, расположенных в продолговатом мозге. По волокнам пути идут импульсы, поддерживающие равновесие и осуществляющие координацию движения.
Боковой канатик спинного мозга содержит следующие проводящие пути:
1) задний спинно-мозжечковый тАУ несет проприоцептивные импульсы в мозжечок;
2) передний спинно-мозжечковый тАУ идет в кору мозжечка;
3) латеральный спинно-таламический тАУ проводит импульсы болевой и температурной чувствительности;
4) латеральный корково-спинномозговой (пирамидный) тАУ проводит двигательные импульсы от коры большого мозга к спинному мозгу;
5) красноядерно-спинномозговой тАУ проводит импульсы автоматического (подсознательного) управления движениями и поддерживает тонус скелетных мышц.
Задний канатик содержит пути сознательной проприоцептивной чувствительности (сознательное суставно-мышечное чувство), которые направляются в головной мозг и корковый конец двигательного анализатора, передают информацию о состоянии мышц тела, его частей в пространстве. На уровне шейных и верхних грудных сегментов спинного мозга задние канатики промежуточной бороздой делятся на два пучка тАУ Тонкий пучок Голля и клиновидный пучок Бурдаха.
Рис.7
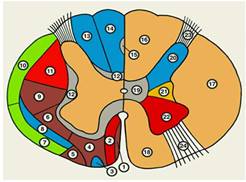
Схематическое изображение поперечного разреза спинного мозга.
Слева обозначены проводящие пути, справа тАФ участки серого вещества; одинаковыми цветами обозначены группы проводящих путей и соответствующие им участки серого вещества; синим цветом тАФ чувствительные пути и задний рог, красным тАФ пирамидные пути и передний рог, серым тАФ собственные пучки спинного мозга и промежуточное вещество, зеленым тАФ восходящие пути экстрапирамидной системы, желтым тАФ боковой рог: 1 тАФ покрышечно-спинномозговой путь; 2 тАФ передний корково-спинномозговой путь; 3 тАФ передний спиноталамический путь; 4 тАФ преддверно-спинномозговой путь; 5 тАФ оливоспинномозговой путь; 6 тАФ ретикул-спинномозговой путь: 7 тАФ передний спиномозжечковый путь; 8 тАФ латеральный спиноталамический путь; 9 тАФ красноядерно-спинномозговой путь; 10 тАФ задний спиномозжечковый путь; 11 тАФ латеральный корково-спинномозговой путь; 12 тАФ собственные пучки спинного мозга; 13 тАФ клиновидный пучок; 14 тАФ тонкий пучок; 15 тАФ овальный пучок; 16 тАФ задний канатик; 17 тАФ боковой канатик; 18 тАФ передний канатик: 19 тАФ промежуточное вещество; 20 тАФ задний рог; 21 тАФ боковой рог; 22 тАФ передний рог; 23 тАФ задний корешок; 24 тАФ передний корешок.
4. Головной мозг
Головной мозг, передний отдел центральной нервной системы позвоночных животных и человека, помещающийся в полости черепа. Головной мозг тАУ материальный субстрат высшей нервной деятельности и главный регуляторвсех жизненных функций организма. В целом головной мозг состоит из скоплений тел нервных клеток
,
нервных трактов и
кровеносных сосудов
. Нервные тракты образуют белое вещество мозга и состоят из пучков
нервных волокон
, проводящих импульсы к различным участкам серого вещества мозга - ядрам или центрам - или от них. Проводящие пути связывают между собой различные ядра, а так же головной мозг со
спинным мозгом
.
При осмотре препарата головного мозга хорошо заметны три его наиболее крупные составные части. Это парные полушария большого мозга, мозжечок и мозговой ствол.
В функциональном отношении мозг можно разделить на несколько отделов:
1)
передний мозг, состоящий из
конечного мозга
(больших полушарий) и
промежуточного мозга,
2)средний мозг
,
включающий ножки мозга и четверохолмие,
3)задний мозг, состоящий из
мозжечка
и
варолиева моста
,
4)продолговатый мозг
.
Продолговатый мозг, варолиев мост и средний мозг вместе называются
стволом головного мозга.
4.1 Продолговатый мозг
Продолговатый мозг находится между задним и спинным мозгом. Нижняя граница продолговатого мозга соответствует уровню большого затылочного отверстия, или месту выхода корешков I пары спинномозговых нервов, верхняя граница проходит по заднему краю моста. Длина продолговатого мозга взрослого человека составляет в среднем 25 мм. Верхняя часть продолговатого мозга в отличие от нижней имеет некоторое утолщение, чем напоминает форму конуса. Борозды мозга являются продолжением борозд спинного мозга. По сторонам от передней срединной щели на вентральной поверхности продолговатого мозга расположены выпуклые, постепенно сужающиеся внизу пирамиды, образованные пирамидными трактами, часть волокон которых формирует перекрест пирамид. Латеральнее пирамиды с двух сторон находятся возвышения тАУ оливы, отделенные от пирамиды передней латеральной бороздой, из которой выходят корешки подъязычного нерва (XII пара черепных нервов). В нижней части дорсальной поверхности продолговатого мозга проходит дорсальная срединная борозда, по сторонам которой заканчиваются утолщениями тонкий и клиновидный пучки задних канатиков спинного мозга, отделенные друг от друга задней промежуточной бороздой. В утолщениях пучков располагаются соответствующие ядра, от которых отходят волокна, формирующие медиальную петлю. Последняя на уровне продолговатого мозга образует перекрест. Пучки этого перекреста располагаются в межоливном слое, дорсальнее пирамид. Из заднелатеральной борозды продолговатого мозга выходят тонкие корешки языкоглоточного (IX пара), блуждающего (X пара) и добавочного (XI пара) черепных нервов, ядра которых лежат в дорсолатеральных отделах продолговатого мозга. На дорсальной поверхности части бокового канатика расширяются и вместе с волокнами от клиновидного и нижнего ядер образуют нижние мозжечковые ножки, ограничивающие снизу ромбовидную ямку; верхняя часть дорсальной поверхности участвует в образовании дна IV желудочка. Серое вещество продолговатого мозга представлено скоплениями нейронов, которые образуют нижние оливные ядра. Дорсальнее пирамид находится ретикулярная формация, состоящая из переплетений волокон тАУ нервных клеток.
Рис.8
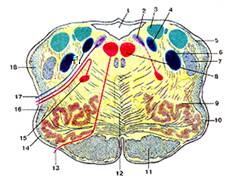
Продолговатый мозг. Поперечный разрез на уровне олив.
1-четвертый желудочек; 2-дорсальное ядро блуждающего нерва; 3-ядро вестибулярною нерва; 4-ядро одиночного пути; 5-задний (дорсальный) спинно-мозжечковый путь; 6-спинномозговое ядро тройничною нерва; 7-спинно-мозговой путь тройничного нерва; 8-ядро подъязычного нерва; 9-оливное ядро; 10-олива; 11-корково-спинномозговой путь (пирамидный); 12-медиальная петля; 13-подъязычный нерв; 14-передние наружные дуговые волокна; 15-двойное ядро; 16-спинно-таламический и спинно-покрышечный пути; 17-блуждающий нерв; 18-вентральпый (передний) спинно-мозжечковый путь.
Продолговатый мозг осуществляет рефлекторную и проводниковую функции. По чувствительным волокнам корешков черепных нервов он получает информацию (импульсы) от кожи, слизистых оболочек и органов головы, а также от рецепторов гортани, трахеи, внутренних органов грудной клетки (легкие, сердце), пищеварительной системы. Через продолговатый мозг осуществляются многие простые и сложные рефлексы. Например: 1) защитные тАУ кашель, чиханье, рвота, слезоотделение, мигание; 2) пищевые тАУ сосание, глотание, отделение пищеварительного сока; 3) сердечно-сосудистые, регулирующие деятельность сердца и кровеносных сосудов; 4) автоматически регулируемый дыхательный центр, обеспечивающий вентиляцию легких; 5) вестибулярные ядра, участвующие в осуществлении рефлексов позы, в перераспределении тонуса мышц.
Кроме того, через продолговатый мозг проходят пути, которые соединяют двусторонней связью кору головного мозга, промежуточный и средний мозг, мозжечок и спинной мозг.
4.2 Задний мозг
Задний мозг состоит из
мозжечка
и варолиева моста.
Мозжечок располагается кзади от моста и от верхней части продолговатого мозга, заполняя большую часть задней черепной ямки. В мозжечке различают верхнюю и нижнюю поверхности, границей между ними является задний край мозжечка, где проходит глубокая горизонтальная щель. Мозжечок лежит в дорсальной поверхности ствола мозга, охватывает его с боков и при помощи ножек соединяется с его частями: верхние мозжечковые ножки соединяют мозжечок со средним мозгом, средние- с мостом; нижние тАУ с продолговатым мозгом. В мозжечке различают два полушария и непарную среднюю часть тАУ червь мозжечка. На верхней и нижней поверхности полушарий и червя находится много параллельно идущих щелей мозжечка, между которыми находятся длинные и узкие извилины мозжечка.
Рис.9
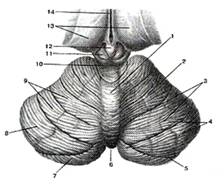
Мозжечок. Вид сверху. 1-червь мозжечка; 2-полушарие мозжечка; 3-щели (борозды) мозжечка; 4-листки мозжечка; 5-горизонтальная шель; 6-задняя вырезка мозжечка; 7-нижняя полулунная долька; 8-верхняя полулунная долька; 9-четырехугольная долька; 10-нижние холмики крыши среднего мозга; 11-верхний холмик
Вместе с этим смотрят:
Бодрствование - нейрофизиологический процесс человека
Генетична iнформацiя
Естествознание в системе наук
Загальна характеристика i особливостi життiдiяльностi бактерiй родини Enterobacteriaceae
Изменчивость организмов