Бiологiчний контроль збудникiв хвороб рослин
ЗмiстВСТУП
РОЗДРЖЛ 1. Сучасний стан епiдемiологii вiрусiв вищих рослин
РОЗДРЖЛ 2. Шляхи розповсюдження вiрусiв рослин в природi та роль факторiв навколишнього середовища
2.1 Клiмат та погоднi умови
2.2 Розповсюдженiсть вiрусiв. Бiотичнi фактори
2.3 Розповсюдженiсть вiрусiв. Абiотичнi фактори
2.4 Шляхи передачi вiрусiв рослин в природних умовах
Роздiл 3. Найбiльш поширенi вiруснi захворювання серед вищих рослин
3.1 Кiлькiсна характеристика та класифiкацiя вiрусiв рослин
3.2 Вiруси, якi вражають певнi види рослин
3.3 Вiруси, якi уражують широке коло рослин господарiв
3.4 Боротьба iз вiрусними хворобами рослин
РОЗДРЖЛ 4. Бiологiчний метод: його суть та використання бiопрепаратiв
4.1 Бiологiчний метод
4.2 Ефективне застосування бiопрепаратiв при вирощуваннi овочевих культур
4.3 Бактерiальнi препарати i бiогумат та iхнi властивостi
ПРАКТИЧНА ЧАСТИНА
Аналiз способiв застосування бактерiальних препаратiв i регулятора росту бiогумату на овочевих культурах
Ефективнiсть використання бактерiальних препаратiв i регулятора росту бiогумату при вирощуваннi овочевих культур
Застосування бактерiальних препаратiв i регулятора росту бiогумату на помiдорах
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОРЗ ЛРЖТЕРАТУРИ
ВступАктуальнiсть. Протягом останнього часу в фiтовiрусологи, як i в iнших молекулярно-бiологiчних дисциплiнах, спостерiгаiться значний iнтерес до розвитку дослiджень на популяцiйному та еколого-популяцiйному рiвнi, i ця тенденцiя i загальною для багатьох традицiйних молекулярно-бiологiчних дослiджень. Значнi успiхи у фiтовiрусологи на молекулярно-бiологiчному рiвнi, такi як розшифровка структури генiв, бiлок-бiлковоi та бiлок-НК взаiмодii, розшифровка АК-послiдовностей та тривимiрноi будови структурних та регуляторних вiрусних бiлкiв та багато iншого контрастуi з нашими знаннями вiдносно поведiнки вiрусiв рослин в природнiх умовах бiоцеозiв. Поява в останнi десятирiччя нового поколiння дiагностичних методiв для визначення наявностi фiтовiрусiв (таких як РЖФА, RIA, RIPA, ПЛР, ЛЛР, та iн) дали змогу на новому рiвнi вивчати розповсюдженiсть фiтовiрусiв в навколишньому середовищi. Актуальнiсть даного питання полягаi в його як фундаментальностi, так i в практичному значеннi. Визначення ареалу розповсюдженостi, механiзмiв передачi, кола рослин-господарiв, реакцii на змiни оточуючого середовища даi можливiсть не тiльки бiльш повно характеризувати того чи iншого представника царства Vira, але i дасть змогу прогнозувати появу та розвиток вiрусних хвороб i правильно виробляти стратегiю i тактику боротьби з ними тАФ запровадження стiйких сортiв, цiлеспрямована боротьба з резерватами та переносниками, отримання безвiрусного посадкового матерiалу.
Перспективним напрямком i моделювання розвитку хвороб, однак для вiрусних хвороб рослин адекватнi моделi практично вiдсутнi, на вiдмiну вiд фiтопатогенних грибiв та мiкроорганiзмiв.
Встановлення можливих взаiмозв'язкiв мiж наявнiстю вiрусних антигенiв в рослинах та ТСрунтi i дуже важливим у розумiннi розповсюдження фiтовiрусiв рослин та циркуляцii iх в агроценозах.
Предмет дослiдження тАУ збудники хвороб рослин.
Об`iкт дослiдження тАУ застосування бiологiчного контролю збудникiв хвороб рослин.
Мета роботи тАУ з`ясувати шляхи проникнення вiрусних iнфекцiй в рослиннi органiзми
Основнi завдання:
1) охарактеризувати сучасний стан епiдемiологii вiрусних захворювань рослин на Украiнi;
2) розглянути шляхи розповсюдження вiрусiв рослин в природi та роль екофакторiв у цьому процесi;
3) дати стислу характеристику найбiльш поширених в Украiнi вiрусiв рослин;
4) розглянути основнi заходи боротьби з вiрусами рослин;
5) проаналiзувати суть та перспективи застосування бiологiчного контролю збудникiв рослин.
Роздiл 1. Сучасний стан епiдемiологii вiрусiв вищих рослинНа порозi двадцять першого столiття епiдемii хвороб рослин все-таки призводять до значних втрат у продовольствi, промисловостi, лiсових та сiльськогосподарських культурах всюди у свiтi. Населення нашоi планети продовжуi збiльшуватись i якщо ми хочемо мати достатньо продовольства, одягу та помешкань для поточного та майбутнiх поколiнь, важливим i зменшення або й зовсiм усунення втрат вiд хвороб рослин.
Епiдемiологiя хвороб рослин являi собою вивчення просторових та часових змiн, що вiдбуваються пiд час епiдемiй хвороб рослин, якi викликанi популяцiiю патогенiв у популяцii рослин. Епiдемiя трапляiться, коли вiдбуваються змiни у iнтенсивностi захворюваностi серед популяцii господаря у часi та просторi. Патогеном можуть бути гриби, бактерii, мiкоплазмоподiбнi органiзми (МПО), вiруси, вiроiди, нематоди або паразитичнi рослини. Рослини можуть бути частиною будь-якоi сiльськогосподарськоi, садовоi, лiсовоi, або природноi екосистеми. Взаiмодiя патогена та рослини-господаря, що призводить до розвитку хвороби, може трапитися у будь-якому наземному, водному, або штучно створеному середовищi. Сума усiх патогенiв, господарiв та середовищних компонентiв являi собою рослинну патосистему.
Наука, що вивчаi епiдемii хвороб рослин (тобто, епiдемiологiя) маi як описову, так i кiлькiсну фази. Спочатку ставиться питання, наприклад, де епiдемii трапляються, коли можуть епiдемii очiкуватися, якi фактори вiдiграють роль у розвитку хвороби. Коли знання патосистеми збiльшуiться, починаються питання бiльш кiлькiсноi природи, наприклад, скiльки iнфекцiйних одиниць патогену потребуiться, щоб започаткувати епiдемiю, скiльки хвороба триваi, як швидко буде хвороба розвиватися, та як далеко може iнфекцiйна одиниця патогену розповсюдитись. Щоб вiдповiсти на цi питання, використовуiться експериментальна та аналiтична технiка класичноi фiтопатологii та технiки, розробленi у агрономii, ботанiцi, хiмii, екологii, ентомологii, генетицi, математицi, метеорологii, фiзицi та статистицi.
Вiруси та мiкоплазмоподiбнi органiзми (МПО) рiдко i окремими, незалежними iнфекцiйними одиницями в оточуючому середовищi. Навiть частки цих патогенiв звичайно пов'язанi з рослиною-господарем або з переносниками (комахами чи нематодами). Таким чином, визначення рiвня iнфекцiйних одиниць в першу чергу робиться шляхом оцiнки титру патогена в рослинах-господарях, або шляхом монiторингу популяцii вiдомих переносникiв та визначення частки популяцii переносника, що несе патоген рослин. У випадку титру вiрусу точну кiлькiсть його не вимiрюють, тодi як комаха або нема-iндii, кожна з яких i переносником патогену, насправдi i iнфекцiйною одиницею або пропагулою епiдемiчного розвитку.
Зiбранi зразки з iнфекцiйними збудниками вiрусноi природи можна iдентифiкувати за допомогою електронного мiкроскопу, вiзуального мiкроскопiчного спостереження або шляхом бiологiчного тестування. Бiохiмiчнi технологii можуть бути корисними для вiрусiв та МПО. При виборi методу монiторингу потрiбно приймати до уваги складнiсть iдентифiкацii, необхiдний час та вартiсть витратних матерiалiв.
Концепцiя збудника не зовсiм чiтка для вiрусiв та МПО. Хоча iснують окремi частки або одноклiтиннi органiзми, вони мало впливають на епiдемiологiю хвороб, спричинених вiрусами або МПО. Для багатьох з цих хвороб патоген передаiться з рослини на рослину за допомогою дiяльностi комах. Деякi ТСрунтовi вiруси переносять нематоди або гриби. Обчислення кiлькостi iнфекцiйних одиниць поза рослиною-господарем роблять за допомогою збирання зразкiв векторiв. Обчислення вiрусiв та МПО в iхнiх рослинах-господарях для епiдемiологiчних завдань даi кращi результати при оцiнцi iнтенсивностi хвороби.
Хоча рослиннi патогени переносять рiзнi ряди комах, найбiльш численнi вектори маi ряд Homoptera. Родини цього ряду включають горбаток, бiлокрилок, кокцид та попелиць. Взагалi, комахи Homoptera переносять 241 вiрус та 45 МПО. Як i у випадку з повiтряними грибами, комах можливо обчислити за iхнiм накопиченням (зiткненням) iз стацiонарними пастками, або iх концентрацii в повiтрi за допомогою всмоктувальних пасток. Далi, можливо визначити кiлькiсть особин, що несуть патоген та здатнi до його трансмiсii.
Оцiнка хвороб рослин i одним з найбiльш важливих та часто найбiльш складних завдань у епiдемiологii хвороб рослин. Оцiнка хвороби на даний час i пiдвалиною статистичного аналiзу даних, зусиль з моделювання та iнтерпретацii патосистеми. Похибка, введена пiд час оцiнки хвороби, може бути дуже малою на початку, але збiльшитись при подальших епiдемiологiчних аналiзах. Отже, ступiнь впевненостi у висновках та спроможнiсть розрiзнювати справжнi ефекти не буде бiльшим, нiж рiвень надiйностi при оцiнцi хвороби.
Оцiнка хвороби звичайно триваi багато часу i тому вiдносно дорога. Кропiтке планування тАФ як, коли, де та ким тАФ хворобу буде оцiнено, дуже iстотне. Планування вимагаi ясноi концепцii мети специфiчного дослiдження та докладне знання патосистеми.
Оцiнки хвороби можуть бути кiлькiсними, якiсними, або комбiнацiiю цих двох типiв. Кiлькiсть наявноi хвороби можна назвати поширенiстю хвороби. До iнтенсивностi хвороби входять розповсюдженiсть хвороби та суворiсть хвороби. Додатково, такi характеристики, як концентрацiя або титр пропагативних або iнфекцiйних одиниць (вiрiонiв) у тканинах, можуть вiддзеркалити iнтенсивнiсть хвороби. Якiснi оцiнки хвороби можуть включати як ринкову якiсть (вмiст протеiну, олii, або вмiст поживних речовин у господарi або Його продуктi) так i такi фактори, як поживнiсть, смак та запах.
Поширенiсть хвороби тАФ це число одиниць рослин, що вiзуально уражено, звичайно до загального числа одиниць, що оцiнено. Якщо розглядати рослину в цiлому як одиницю, тодi поширенiсть хвороби i пропорцiя або процент рослин iз симптомами хвороби. Поширенiсть хвороби може також бути оцiнена, використовуючи окремi гiлки, листки або корiння як рослинну одиницю.
Поширенiсть та суворiсть як вимiри оцiнки вживають разом, i правомiрнiсть кожного вимiру повинна бути встановлена для кожноi окремоi патосистеми. Поширенiсть часто найбiльш легка та найбiльш швидка мiра хвороби. Число зiв'ялих рослин томату, почорнiлi колосся пшеницi, корiнь iз пухлинами або листя з некрозами чи мозаiчними симптомами можуть бути порахованi досить точно та акуратно. Звичайно поширенiсть i вiдповiдною мiрою хвороби для патосистем, у котрих один некроз на одиницю рослини i фатальним, також як i у випадку з хворобами в'янення та багатьма системними вiрусними хворобами. Поширенiсть симптоматичних листкiв aбo гiлок може також бути мiрою хвороби для комерцiйних плодових дерев, тiньових лiсiв, тощо. Оцiнка або вимiрювання суворостi хвороби часто бiльш тяжке та вiдбираi бiльше часу. Також, точнiсть та вiдтворюванiсть може бути меншою, нiж у випадку поширеностi. Суворiсть, однак, може бути бiльш важливою та корисною мiрою дослiдження хвороби для багатьох патосистем, що мають вiдношення до управлiння хворобами, динамiки чисельностi та оцiнки втрат врожаiв.
У бiльшостi випадкiв, суворiсть хвороби розглядають як частку розглянутих рослинних одиницiь. Свiдчення з фiзiологii рослин та дослiджень втрат врожаiв показують, що врожай рослин може бути визначений iз загальноi суми уражених областей або, бiльш точно, iз загальноi суми здорових областей. Це примушуi епiдемiологiв винайти шляхи для оцiнки ступеню абсолютноi суворостi хвороби, хоча необхiдно оцiнити втрати врожаю. Однак на сьогоднiшнiй день можуть бути застосованi декiлька рiзноманiтних шляхiв репрезентацii iнтенсивностi.
Термiни та частоти оцiнок хвороби визначаються пагосистемою та метою оцiнки. Оцiнки можуть ТСрунтуватися на календарних строках, фiзiологiчно-середовищних чинниках, або в залежностi вiд стадiй росту. Календарна шкала i найбiльш часто використаною шкалою часу у епiдемiологiчних дослiдженнях, що часто проводять через 5-, 7-, 10-, або 30-деннi iнтервали. Календарнi оцiнки i зручними та дозволяють скласти регулярний розклад роботи. Такi оцiнки також забезпечують регулярно розмежованi данi для аналiзу прогресу хвороби, у якому час використаний як незалежна змiнна.
Фiзiологiчно-середовищна шкала часу використовуi середовищнi властивостi або комбiнацii властивостей, що i важливими для фiзiологiчного стану господаря або патогену. Цей часовий масштаб ТСрунтуiться на концепцii, що певне число середовищних одиниць повинне бути акумульовано для того, щоб ключова фiзiологiчна подiя трапилася. Число днiв з певною температурою, вищою за мiнiмальну тАФ приклад цього типу шкали часу. Цей тип шкали вимагаi безперервного монiторингу змiнних середовища (або доступ до таких даних). Складання розкладу у такому випадку оцiнки дещо бiльш важке, нiж з календарною шкалою, якщо бажанi регулярнi iнтервали одиниць. Такими шкалами користувались вiдносно рiдко в фiтопатологii; однак, приклади i, та й подальше дослiдження переваг цiii шкали часу виправдане.
Розширення фiзiологiчно-середовищноi шкали базуiться на стадiях росту господаря. В межах цiii пов'язаноi iз господарем шкали, рослина сама слугуi як iнтегратор всiх середовищних ефектiв. Припускаючи, що стадii росту ТСрунтуються на ключових подiях у життiвому циклi господаря, оцiнки хвороби при цих ключових стадiях забезпечують iнформацiю, що може бути корисною для аналiтичноi або модельноi мети, для оцiнки втрат врожаю, або для оцiнки матерiалу у програмах вирощування рослин, стiйких до хвороб.
Частота оцiнок може залежати вiд частоти подiй, вимог щодо iнформацii про рiзнi стадii епiдемii, або економiчних мiркувань. На фiзiологiчно-середовищнiй або шкалi стадiй росту, частота специфiчних подiй (наприклад, акумуляцiя температурних днiв або перехiд до наступноi стадii росту) визначить частоту оцiнок хвороби. Якщо потрiбен докладний статистичний або математичний аналiз епiдемii, частота оцiнки може визначатись потребою в специфiчнiй кiлькостi спостережень або потребi в спостереженнях пiд час "критичних" частин епiдемii. Для епiдемii, що триваi 90 днiв, тижневi оцiнки дадуть 12-13 рiвномiрно розподiлених спостережень. Ця кiлькiсть певно i достатньою для бiльшостi випадкiв; однак, якщо економiчнi мiркування диктують, що можна зробити менше нiж 12 спостережень, буде бiльш сприятливим зробити оцiнки з нерегулярними iнтервалами.
Економiчнi мiркування при оцiнцi хвороби повиннi бути врiвноваженi з потребою у специфiчнiй iнформацii. Якщо час i обмежувальним фактором (як це i i звичайно), тодi необхiдно визначити мiнiмальне число оцiнок. Мета оцiнки та знання джерел змiн в межах та серед експериментальних одиниць буде диктувати, чи краще зробити бiльше спостережень у одному мiсцi, або менше спостережень у багатьох мiсцях.
Можна зробити висновок про те, що маючи досить ТСрунтовну теоретичну базу, ми ще не маiмо бiльш-менш стандартних методик щодо об'iктивноi оцiнки розвитку вiрусних iнфекцiй в агроценозах. Цей факт свiдчить про актуальность цiii проблеми як ii вузькому (вiрусологiчному) так i в широкому (сiльськогосподарському та екологiчному) розумiннях.
Роздiл 2. Шляхи розповсюдження вiрусiв рослин в природi та роль факторiв навколишнього середовищаВiруси рослин мають здатнiсть досить швидко розповсюджуватися в бiоценозах. Вони можуть поширюватися завдяки переносникам (клiщi, попелицi, нематоди, гриби), передаватися при вегетативному розмноженнi через бульби, живцi, цибулини. Бiльшiсть фiтовiрусiв передаються щепленим. Близько 90 рiзних вiрусiв рослин передаються насiнням, деякi фiтовiруси можуть викликати iнфекцiю без участi переносникiв, коли збудник знаходиться в грунтi. Знанiй екологii збудника необхiдне для використання необхiдних засобiв боротьбi проти нього (рис. 1.).
При ураженнi рослин тАФ господарiв фiтовiрусами найчастiше розвиваiться системна реакцiя. При цьому iнфекцiя часто протiкаi з яскраво вираженими симптомами у виглядi мозаiки, скручування, хлорозу, появи енацiй та iн. Така iнфекцiя може закiнчуватись навiть загибеллю iнфiкованих частин рослин або i всiii рослини. Разом з тим, iнколи перебiг iнфекцiйного процесу проходить без чiтких ознак хвороби, якi помiтнi за габiтусом рослин. Часто уражений органiзм може бути прихованим носiiм iнфекцii. Продуктивнiсть такi(Х рослин знижуiться в незначнiй мiрi. Так реагують на вiрусну iнфекцiю рослини-резерватори, переважна бiльшiсть яких вiдноситься до бур'янiв.
Бур'яни (щириця бiла, гiрчиця бiла, сурiпиця звичайна, пирiй повзучий та iн.) займають особливе мiсце серед факторiв, що знижують врожай сiльськогосподарських культур. Для цих рослий характернi велика енергiя насiннiвого та вегетативноi о розмноження, тривале зберiгання схожостi насiння. Вони сильно виснажують та висушують грунти, часто являються джерелом розповсюдження хвороб та шкiдникiв сiльськогосподарських культур. Джерелом вiрусiв служить також насiння бур'янiв. Завдяки його достиганню та обсипанню до збору сiльськогосподарських культур, насiння накопичуiться на полях у великiй кiлькостi. Так, в одному шарi товщиною 30 см знаходиться в середньому вiд 100 млн. до 4 млрд. i бiльше насiння бур'янiв на 1 га. При обробцi грунтiв значна iх частина перемiщуiться в верхнiй горизонт, i iх кiлькiсть тут в сотнi i тисячi разiв перевищуi кiлькiсть насiння, що висiваiться. Крiм того, грунт може бути забрудненшiй кореневищами бур'янiв, що також можуть бути ураженi вiрусами.
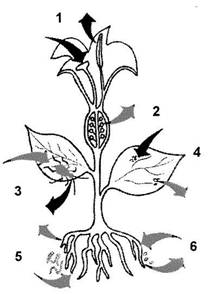
Рис. 1. Основнi шляхи передачi фiтовiрусiв.
1 - передача за допомогою пилку; 2 - передача за допомогою насiння; 3 - за допомогою векторiв-комах; 4 - при прямому контактi з рослинами (механiчному пошкодженнi); 5 - за допомогою еекпюрiв-нематод; 6 - за допомогою векторiв-грибiв.
Так, вiрус мозаiки цукрового буряку уражуi також щирицю, осот, кормовi боби. Вiруси, що викликають жовтяницю буряка, можуть перезимовувати в багатьох розповсюджених бур'янах.
Багато вiрусiв швидко мiгрують завдяки переносникам. Переносниками можуть бути попелицi, цикадки, клiщi, нематоди, гриби та iн. Напевно, перше мiсце по передачi вiрусiв серед комах займають попелицi. Такi вiруси, як вiрус огiрковоi мозаiки, вiрус жовтоi карликовостi ячменю, У-вирус картоплi, вiрус мозаiки цукрового буряку, вiрус жовтяницi цукрового буряку можуть передаватися рiзними видами попелиць. В деяких випадках, наприклад, мiж вiрусом жовтоi карликовостi ячменю та попелицями, iснуi симбiотичний взаiмозв'язок. При живленнi попелиць на уражених цим вiрусом рослинах пiдвищуiться iх плодовитiсть. Активними переносниками вiрусу смугастоi мозаiки пшеницi i клiщi Aceria tulipae Keit та Асегiа tritici Schev. ВСМП може довгий час циркулювати в природi. Зимуi вiрус в iнфiкованих рослинах i клiщах. З весни та до збору врожаю клiщi iнтенсивно розмножуються на рослинах, а потiм мiгрують на бур'яни (пирiй повзучий, стоколос безостий та iн) i кукурудзу. Восени вошi мiгрують на сходи озимих та уражують iх вiрусом (40).
Попелицi i клiщi, як i iншi летючi комахи, i одними з важливих факторiв, що визначають дальнiсть розповсюдження вiруса вiд джерела. Так, за один день попелицi вiдносяться вiтром на десятки i навiть сотнi кiлометрiв.
Грунт i також одним iз джерел iнфiкування рослин вiрусами. Потрапляючи в грунт, вiруси здатнi iнфiкувати новi рослини. РЖснуi три групи вiрусiв, що передаються через грунт:
1. вiруси, що передаються грибами;
2. вiруси, що передаються нематодами;
3. вiруси, переносник яких невiдомий.
Крiм вiрусiв, що переносяться або нематодами з родiв Longidorus, Xiphinema i Trichodorus, або зооспорами ТСрунтових грибiв з родiв Polymixa i Olpidium, iснуi група ТСрунтових вiрусiв, представники якоi "вiльними" вiбрiонами зустрiчаються в грунтi i водi та можуть переходити без допомоги переносникiв вiд хворих до здорових рослин (non-vectored viruses), хоча деякi з них можуть переноситися нематодами або грибами.
Незважаючи на свою рiзноманiтнiсть, грунтовi вiруси мають ряд спiльних ознак (за винятком ВОМ):
- не розповсюджуються лiтаючими переносниками;
- накопчуються в рослинах-господарях у високих концентрацiях i видiляються через коренi iнфiкованих рослин в грунт;
- надзвичайно стiйкi поза живою клiтиною;
- можуть iнфiкувати здоровi рослини через коренi без допомоги переносникiв;
- мають, як правило, широке коло рослин-господарiв;
- бiльшiсть з них мають простий однокомпонентний геном;
- деякi з них можуть розповсюджуватись за допомогою безпосереднього контакту.
В грунт вiруси рослин можуть потрапити як прямим (з листових, стеблових i кореневих залишкiв уражених рослин, та при вiддачi вiрусiв коренями iнфiкованих рослин), так i непрямим шляхом (вiрусовмiснi екскременти та вимивання вiрусiв з контамiнованих ТСрунтiв дощовими водами). Так, вiрус огiрковоi мозаiки може проходити через травний канал людини або тварин, а потiм зi стiчними водами потрапляти в грунт i вражати рослини.
Джерелами контамiнацii "вiльних" вiд вiрусiв ТСрунтiв або iнших субстратiв може бути, ймовiрно, невелика кiлькiсть рослин, що постiйно видiляють через коренi вiрус в навколишнi середовище. Деякi вiруси настiльки призвичилися до цього способу розповсюдження, що iнфiкують тiльки кореневу систему рослин-господарiв або окремi частини iх надземних органiв. Дiагностика в такому випадку ускладнена, i можна припустити, що вiруси цiii групи розповсюдженi бiльш широко, нiж думали до останнього часу.
На даний перiод в лiтературi описанi 11 вiрусiв, у яких встановлена вiддача вiрусiв через коренi i 11 вiрусiв, у яких показана iнфекцiя рослин через коренi без переносникiв.
Численнi вiруси рослин мiстяться в ТСрунтi i в поверхневiй водi, особливо в сiльськогосподарських областях. Ступiнь адсорбцii i тривалiсть перiоду, на протязi якого iнфекцiйнiсть зберiгаiться i який може тривати тижнi, а iнодi i мiсяцi, залежить вiд захисту вiрусу i факторiв зовнiшнього середовища. Серед останнiх це тип ТСрунту, рiвень вологостi, рН, температура, вмiст органiчних субстанцiй i звичайно зв'язана з цим бiологiчна активнiсть середовища. Багато вiрусiв стабiльнi i здатнi iнфiкувати рослини без допомоги векторiв, а саме нематод та розповсюджених в ТСрунтi зооспор грибiв. Передача вiрусу з ТСрунту та води ймовiрно можлива через непошкодженi коренi рослин. Вiруси, якi i в струмках, можуть переноситись на великi вiдстанi. РЖнодi новi вiруси, не знайденi у рослинах-господарях, були iзольованi з води та ТСрунту.
До теперiшньоi о часу приблизно 9 вiрусiв були iдентифiкованi i кiлька iнших iзольованi з ТСрунту i води лiсових екосистем. Вони головним чином належать до: Potex,- Tombus-, Tobamo-, Poty- та Necrovirus груп. Крiм цього, цi вiруси та принаймнi 7 представникiв iнших таксономiчних груп, особливо Nepo- та Ilarviruses, були знайденi в деревовидних лiсових рослинах. Такi збудники вiрусних iнфекцiй можуть бути бiльш або менш шкiдливi для iнших видiв рослин. Також багато вiрусiв було iдентифiковано в квiткових лiсових рослинах. Цi патогени являють собою потенцiйну загрозу для економiки лiсового господарства.
Невелика кiлькiсть дослiджень поширення вiрусiв в цих субстратах (ТСрунтi та водi) спонукаi нас проводити роботу по iдентифiкацii та розповсюдженостi вiрусних iнфекцiй в агроценозах Украiни.
Останнiм часом увага дослiдникiв була привернута до вiрусних систем збереження, як видно з екологiчних робiт. Для деяких вiрусiв добре розвинена здатнiсть до збереження на полi (дiльницi) компенсуiться обмеженою здатнiстю до поширення на новi дiльницi. Навпаки, iншi вiруси, особливо деякi з тих, що передаються персистентим способом повiтряними векторами, широко передаються на новi дiльницi, але не здатнi до довгого збереження на данiй дiльницi (якщо мова йде виключно про агроценоз). Володiння ефективними способами як передачi, так i збереження, i характерною рисою деяких з бiльшостi постiйно циркулюючих вiрусiв.
Тому що вiруси з однiii таксономiчноi групи мають подiбну систему виживання, а рiзнi типи рослинних угрупувань ймовiрно мають схильнiсть до рiзних видiв систем виживання, можна очiкувати, що специфiчнi групи вiрусiв будуть краще адаптованi до специфiчних рослинних угрупувань. В середовищi, придатному для контактноi передачi тобамо- та потексвiрусiв, рослини виробляють високу концентрацiю вiдносно стабiльних часточок в iхнiх господарях. РЖншi адаптованi до культивованих рослин вiруси (CULPAD viruses), включають iларвiруси, якi головним чином можна знайти в дерев'янистих видах. Вони передаються пилком при запиленнi рослин. Вiруси, якi переносяться за допомогою грибiв (групи вiрусу жовтоi мозаiки ячменю), персистують на рестингспорах iхнiх векторiв i вiддають перевагу монокультурам. З iншого боку, i вiруси, адаптованi до диких рослин (WILPAD viruses) якi включають багато груп, таких як непо-, тобра-, гемiнi-, лютеовiруси. Цi вiруси мають широкий спектр господарiв i довготривалу персистенцiю на iхнiх векторах. Цi властивостi пiдходять до виживання в угрупуваннях, якi складають багато видiв рослин.
Екологiя вiрусiв рослин, а вiднинi i епiдемiологiя вiрусних хвороб рослин не повинна розглядатись як вiдбiр незмiнних зразкiв, вони можуть змiнюватись пiд впливом багатьох факторiв, таких як довгострокова змiна погоди, змiни в методах вирощування рослин, змiни екологiчних параметрiв ценозу. Для прикладу, вирощування рослин з використанням напiвпрозорих пластикових листiв в деяких краiнах привело до поширення вiрусiв на захищених полях, а в Англii зростання замiщення ярого ячменя на озимий супроводжуiться зростанням кiлькостi i поширенням вiрусiв, якi i проблемними для цих культур.
Крiм того, до цих "стороннiх" змiн в методах культивування сiльськогосподарських рослин i бiльш прямi наслiдки вiд спроб контролювати вiруси. Наприклад, вирощування культури томатiв, що мають ген стiйкостi до ВТМ (Тm-1), призводять до селекцii i поширення нестiйких штамiв, але цi штами, очевидно, мають лiмiтовану бiологiчну придатнiсть, i коли тиск, нав'язаний стiйкою культурою, знiмаiться, вони зменшуються в поширеннi.
Найiмовiрнiше, вiруси рослин протидiють стiйкостi, що контролюiться одним чи двома генами господаря, бiльш повiльно, або не контролюються взагалi. Це очевидно вiдбуваiться через те, що iхнi геноми мають обмежену iмнiсть кодiв i iхнiй генний продукт i багатофункцiональним, або через те, що iхнi засоби передачi обмежують ступiнь, до якого рослини з стiйким генотипом пiддаються iнокулюму. В цiй "боротьбi генiв" нестiйкi варiанти можуть з'являтися, але як вважаiться, можуть бути вiдносно невдалими. Протилежними в цiй взаiмодii i селективнi ефекти дii вiрусiв на iхнiх господарiв, якi звичайно проявляються пiсля багатьох рокiв. Таким чином, гени для толерантностi або резистентностi до iнфекцii звичайно знаходять на культивованих посiвах на дiлянках, де рослини постiйно або регулярно пiддаються вiруснiй iнфекцii довгий промiжок часу тАФ резистентнiсть кукурудзи до вiрусу смугастостi кукурудзи в РЖсландii, резистентнiсть картоплi до декiлькох вiрусiв в Пiвденнiй Америцi i тому прикладом.
В епiдемiологii географiчний дiапазон факторiв, якi працюють, i дуже важливим, тому шкала подiй збiльшуiться, що приводить до бiльшоi складностi системи. РД багато прикладiв того, що можна домогтися при детальному вивченнi розповсюдженостi вiрусiв на специфiчних дiлянках. Важкi для розумiння з точки зору епiдемiологii вiрусу великi континентальнi землянi маси, де фактори, дiючi на протязi сотень кiлометрiв, можуть привести до значних ефектiв, i де щорiчна сфера дii iнфекцii може пiддаватися сильним флуктуацiям. В таких областях останнi досягнення методiв, заснованих на вивченнi великомасштабних рухiв повiтряних мас, можуть частково принести успiх для картографii мiграцii повiтряних векторiв. Дiйсно, такий спосiб вивчення впливу однiii частини континенту на iншiй показаний на прикладi впливу великомасштабного перемiщення вiрусу коричневоi плямистостi рису.
Для аналiзу причин i перебiгiв епiдемiй останнiм часом з'явилося багато корисних методик. Велика кiлькiсть рослин може зараз бути протестована на присутнiсть вiрусноi iнфекцii з допомогою чутливих серологiчних методiв, таких як ELISA. Досягнуто прогресу в детекцii вiрусiв серологiчними методами в iндивiдуальних векторних органiзмах, зiбраних на полях, та рiзнi генотипи вiрусiв можуть бути вивченi з допомогою гiбрiдiзацii комплементарноi ДНК. Такою i робота по порiвняльнiй екологii рiзних штамiв вiрусiв. Можна очiкувати, що комп'ютерний аналiз може допомогти розiбратися з великими масивами даних, одержаних пiсля рiзнобiчного вивчення i постiйних записiв стану зовнiшнього середовища, а також допомогти зробити життiво необхiдний висновок по поясненню подiй та iх прогнозуванню.
Ситуацiя, що iснуi в дiйсностi по розвитку кожного вiрусу в конкретнiй мiсцевостi або в свiтовому масштабi, i результатом складноi взаiмодii багатьох фiзичних та бiологiчних факторiв. Розумiння екологii вiрусу щодо окремоi культури та конкретноi мiсцевостi важливе для розробки вiдповiдних методiв боротьби iз захворюваннями, що вiн викликаi. Як i по вiдношенню до бiльшостi iнших облiгатних паразитiв, основними екологiчними факторами, що слiд розглядати, звичайно вважають шляхи передачi вiрусiв вiд рослини до рослини, а також шляхи, якими iншi фактори впливають на розповсюдженiсть вiрусу.
Вiрус, що зберiгаi стабiльнiсть структури, знаходячись як всерединi рослини, так i зовнi i досягаi високих концентрацiй в тканинах, маi, зрозумiло, бiльшу ймовiрнiсть вижити та розповсюдитись, нiж той, що цими властивостями не володii. Наприклад, ВТМ може на протязi довгого перiоду зберiгатись в мертвому рослинному матерiалi, що знаходиться в грунтi, котрий в такому разi стаi джерелом iнфекцii для наступних культур.
Вiруси або штами вiрусiв, що перемiщуються по рослинi повiльно вiд того мiсця, де пройшло ураження, мають меншу ймовiрнiсть вижити i потiм ефективно розповсюджуватись, нiж тi з них, що транспортуються швидко. Швидкiсть руху вiрусiв вiдiграi певну роль, якщо ii розглядати з точки зору тривалостi життя окремих рослин-господарiв. Так, вiруси, що уражують довгоживучi дерева та кущi, можуть рухатись по своiм рослинам-господарям значно повiльнiше, нiж тi, що iнфiкують однорiчнi рослини. Вiруси, що можуть проникати в насiння i зберiгатись там, мають суттiвi переваги порiвняно з iншими, з точки зору розповсюдження, а також здатностi до виживання. Розповсюдження вiрусу некрозу тютюну у бiльшостi рослин-господарiв обмежене кореневою системою, i iснування цього вiрусу в природi залежить вiд здатностi грибiв-переносникiв передавати його через грунт i уражувати таким чином iншi рослини. Ймовiрнiсть виживання вiрусу, що викликаi швидко-прогресуюче системне захворювання, яке призводить до загибелi рослини, значно менша, нiж у тому випадку, коли вiрус викликаi у рослин слабкi або середньоi тяжкостi захворювання, при яких вони продовжують вегетувати та розмножуватись. Цикадки iнфiкуються спочатку штамами вiрусу кучерявостi верхiвки цукрового буряку, що викликають слабкi симптоми на рослинi-господарi. Вiрулентнi штами цього вiрусу викликають загибель рослин до того, як на них закiнчить, розвиток наступне поколiння цикадок. Таким чином, в деяких умовах вiрулентнi штами мають тенденцiю до самознищення. В краiнах з високими температурами влiтку може проходити iнактивацiя вiрусiв in vivo в природних умовах. Так, в деяких районах РЖндii бульби картоплi, що зберiгаються на звичайних складах, можуть звiльнитись вiд вiрусу згортання листкiв, в той час як бульби, що зберiгаються в охолоджених сховищах, залишаються повнiстю iнфекцiйними. Аналогiчним чином рослини суницi звiльнялись вiд вiрусу зморшкуватостi.
Таким чином основнi фактори, що впливають на виживання штаму, такi:
- ефективна передача комахами або iншими засобами;
- бiльш швидке розмноження i бiльший врожай, нiж у конкуруючих штамiв;
- розвиток слабкоi або не надто сильноi форми захворювання. Вiруси широко варiюють по вiдношенню до спектру тих видiв рослин-господарiв, що вони уражують. Так, деякi вiруси, що уражують полуницi, обмеженi родом Fragaria. А вiрус бронзовостi томатiв маi дуже широке коло господарiв, до якого входять як однодольнi, так i дводольнi. Вiдомо, що цей вiрус уражаi рослини 166 видiв, що входять до 34 родин.
Вiруси з дуже вузьким колом рослин-господарiв, ймовiрно, виживають тому, що цi рослини багаторiчнi, або розмножуються вегетативно, або внаслiдок того, що такi вiруси передаються насiнням. Для багатьох нематод та вiрусiв, якi вони переносять, характерний широкий спектр рослин-господарiв, до яких входять i багаторiчнi деревоподiбнi рослини.
Оцiнюючи вiруси рослин в цiлому, вважаiться, що найважливiшим агентом розповсюдження, а таким чином, i виживання вiрусiв i лiтаючi комахи-переносники, що живляться соком рослин, зокрема попелицi. Шлях розповсюдження вiрусу в межах окремоi культури, а також швидкiсть та вiдстань, на яку його може бути перенесено, залежить вiд багатьох факторiв (вiруси, що передаються через повiтряне середовище):
- джерело iнокулюму; iнокулюй може потрапити в культуру зовнi, вiд хворих рослин в самiй культурi, що мали вiрус при передачi насiнням або при вегетативному розмноженнi, а також вiд
бур'янiв та iнших рослин, що знаходяться в цiй культурi;
- кiлькiсть потенцiйно iнфекцiйного iнокулюму;
- природа та modus vivendi переносника (наприклад, щодо попелиць, то маi значення чи це колонii, чи крилатi форми);
- передаiться вiрус за допомогою стилету, i вiн циркулюючим чи розмножуiться в переноснику;
- час, коли переносник стаi активним, вiдносно строкiв вегетацii рослин даноi культури;
- погоднi умови.
Ван Дер Планк ще у 1946 роцi запропонував метод, за допомогою якого можна визначити, чи розповсюджуiться вiрус в межах дiлянки вiд ураженоi рослини, чи вiн потрапляi зовнi. Його засновано на припущеннi, що вiрус, який потрапляi зовнi, буде викликати випадкове ураження рослин, тодi як при його розповсюдженнi вiд хвороi рослини до хвороi, ураженi рослини будуть розташованi групами. Таким чином, оцiнюючи кiлькiсть пар уражених рослин вiдносно очiкуваноi, роблять висновок про походження вiрусу.
Для вiрусiв, що розповсюджуються через грунт швидкiсть розповсюдження залежить вiд:
- типу переносника;
- структури, вологостi, щiльностi i т.п. ТСрунту;
- густини рослин-господарiв;
- клiматичних умов;
- iнших факторiв.
Однак, вiдомi i виключення. Так, ВТМ тАФ один з небагатьох вiрусiв, що в помiтних кiлькостях поширюiться через грунт без допомоги переносникiв.
Окрему групу феноменiв складаi перенесення вiрусiв на великi вiдстанi. Бiльшiсть з таких явищ спонтаннi i здебiльшого не пiдлягають передбаченню. Вони порушують усталену рiвновагу в ценозi, i тому довгостроковi прогнози можливi лише на вiдносно стабiльних системах, оскiльки охоплюють коло тiльки вiдомих характеристик.
На ураженiсть деякоi культури захворюванням вiрусноi етiологii значний вплив мають методи ii вирощування та заходи обробки, що застосовують протягом багатьох сезонiв. Тому будь-який прогноз маi мiстити рекомендацii щодо вдалого сiвообiгу, засобiв обробки, термiнiв висiву та збору рослин. Вплив рiзного роду факторiв, що мають значення у данному випадку, можна розглянути на таких прикладах.
Якщо поряд iз полем озимини знаходиться поле яроi пшеницi, ураженоi вiрусом смугастоi мозаiки, то висiв озимини до збору яровоi пшеницi призводить до великого ступеня ii ураження ВСМП, який переноситься клiщами.
Тип сiвообiгу маi деякий вплив на розповсюдженiсть вiрусiв, що здатнi зимувати в бур'янах або у падалицi попередньоi культури. В деяких культурах велика кiлькiсть спонтанно ростучих рослин, що мiстять вiрус, може вегетувати довгий термiн. Донкастер та Грегорi довели, що для того, щоб позбавитись вiд таких рослин картоплi, необхiдно 5-6 рокiв. А у випадку багаторiчних культур кiлькiсть хворих р
Вместе с этим смотрят:
Анатомическое строение растений
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Анатомия человека
Анатомо-физологические механизмы безопасности и защиты человека от негативного воздействия
Бiологiчне рiзноманiття людських рас