Бiологiя iндивiдуального розвитку
Мiнiстерство Науки i Освiти Украiни
ДНУ
iм. О. Гончара
Контрольна робота
З курсу ВлБiологiя iндивiдуального розвиткуВ»
Виконала
Студентка 3 курсу
Спецiальнiть ВлБiологiяВ»
Будко Г.О.
Дпiпропетровськ
2011р.
План
1. Способи гаструляцii
2. Провiзорнi органи зародка у птахiв
гаструляцiя провiзорний зародок птах
1. Способи гаструляцii
Характер гаструляцii в тваринному царствi надзвичайно сильно варiюi, проте здiйснюiться вона за участю порiвняно нечисленнихВа механiзмiв. При гаструляцii спостерiгаiться поiднання декiлькох основних типiв рухiв.
Ва- iнвагiнацiя
РД укручуванням дiлянки клiтинноi стiнки зародка подiбно до того, як вдавлюiться всередину м'яка стiнка гумового м'яча, коли на нього натискають. При iнвагiнацii механiчна цiлiснiсть стiнки бластули не порушуiться. Впячивание однiii з дiлянок бластодермы всередину здiйснюiться цiлим пластом (мал. 1).
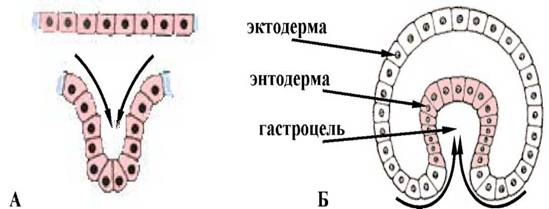
Мал. 1. Схема процесу iнвагiнацii: А - рух пласта клiтин; Б - инвагiнацiйна гаструла.
У ланцетника впячиваються клiтини вегетативного полюса, а у земноводних iнвагiнацiя вiдбуваiться на межi мiж анимальным i вегетативним полюсом в областi сiрого серпа. Процес iнвагiнацii можливий тiльки в яйцях з невеликою або середньою кiлькiстю жовтка. В результатi утворюiться двуслойный мiшок, зовнiшньою стiнкою якого i первинна ектодерма, а внутрiшньою - первинна ентодерма. Бластоцель (первинна порожнина тiла або порожнина Бера) поступово витiсняiться, в ходi впячивання утворюiться гастроцель - первинний кишковик або архентерон, а отвiр за допомогою якого вiн сполучаiться iз зовнiшнiм середовищем, називають бластопором або первинним ротом. Останнiй з усiх бокiв оточений губами бластопора.
Доля бластопора у рiзних тварин неоднакова. У багатьох тварин первинний рот, розвиваючись i диференцiюючись, перетворюiться на рот дефiнiтива дорослого органiзму - первинноротi (черв'яки, молюски i членистоногi). Не менш велика група вторiчноротих, у яких бластопор перетворюiться на анальний отвiр (щетинкочелюстнi, плеченогие, голкошкiрi, кишководишнi) або в нервово-кишковий канал, що знаходиться в задньому кiнцi ембрiона (хордовi).
- iнволюцiя
При цьому вiдбуваiться укручування всередину зародка зовнiшнього пласта клiтин, що збiльшуiться в розмiрах, який поширюiться по внутрiшнiй поверхнi клiтин (мал. 2), що залишаються зовнi.
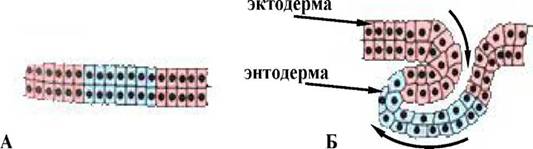
Мал. 2. Схема iнволюцii : А - початковий пласт клiтин, Б - рухи клiтинного пласта в ходi iнволюцii
- iммiграцiя
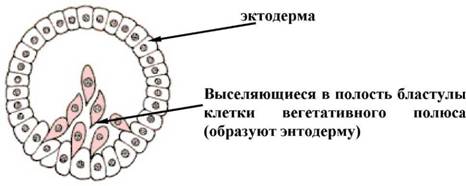
РД найбiльш древнiй спосiб гаструляции, вiдкритий И.И. Мечниковим (1886). РЖммiграцiйна гаструла характерна для багатьох кишковополосних. Вона розвиваiться внаслiдок активного виселення (ингрессии) частини клiтин стiнки бластули всередину бластоцеля. Це виселення клiтин може вiдбуватися як з одного (вегетативного) полюса - унiполярна iммiграцiя, так i з двох протилежних (анимального i вегетативного) полюсiв - бiполярна iммiграцiя (мал. 3).
 _
_
Мал. 3. Унiполярна iммiграцiя (схема)
РЖнодi процеси iммiграцii йдуть без особливого порядку по усiй поверхнi бластули - мультиполярная iммiграцiя. Клiтини, що виселилися, пiзнiше утворюють внутрiшнiй шар гаструли - ентодерму. Таким чином, як i в ходi iнвагiнацii зародок стаi двошаровим. Унiполярна iммiграцiя характерна майже для усiх гiдромедуз, особливо тих, яким властива плаваюча целобластула i мультиполярная iммiграцiя, навпаки, зустрiчаються набагато рiдше. У багатьох кишковополостних, яким властива iммiграцiйна гаструла, вiдбуваiться таке масове, активне виселення клiтин бластодермы, що бластоцель повнiстю заповнюiться iх щiльною масою i зникаi. Важливою особливiстю iммiграцiйноi гаструли i вiдсутнiсть бластопора, а значить немаi i характерного для инвагiнацонной гаструли сполучення гастроцеля iз зовнiшнiм середовищем.
При iммiграцii вiдбуваiться перемiщення окремих клiтин або груп клiтин, не об'iднаних в iдиний пласт. РЖммiграцiя, в тому або iншому видi, зустрiчаiться у усiх зародкiв, але найбiльшою мiрою характерна для другоi фази гаструляции вищих позвонкових (птахiв i ссавцiв).
- делямiнацiя
У тих випадках, коли дроблення закiнчуiться утворенням бластули з невираженою або майже вiдсутньою порожниною (бластоцелем), наприклад морули у деяких кишковополостних, спостерiгаiться розщеплення iдиного клiтинного пласта на два бiльш менш паралельних. Кожна клiтина, що утворюi бластодерму, шляхом мiтотичного дiлення розщепляiться i вiдшнуровуi другий шар (мал. 4). гаструляцii обмежуiться вирiвнюванням внутрiшнiх стiнок клiтин зовнiшнього шару, причому таке вирiвнювання нерiдко йде хвилею вiд однiii сусiдньоi клiтини до iншоi. Уздовж вирiвняних поверхонь формуiться базальна мембрана, що вiддiляi зовнiшнiй клiтинний шар (ектодерму) вiд внутрiшньоi маси клiтин, яка уся стаi ентодермою. Делямiнацiю можна спостерiгати у зародкiв з неповним типом дроблення, таких як плазуни птахи, нижчi ссавцi.
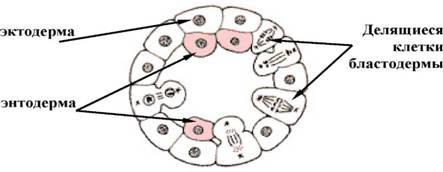
Мал. 4. Деляiинацiя (схема)
тАУ епiболiя
Представляi рух епiтелiальних пластiв клiтин (зазвичай ектодермальних), якi поширюються як одне цiле, а не iндивiдуально i оточують глибокi шари зародка. РЖншими словами, вiдбуваiться обростання дрiбними клiтинами анимального полюса бiльш великих, вiдстаючих в швидкостi дiлення i менш рухливих, клiтин вегетативного полюса (мал. 5).
Формування епiболiчноi гаструли яскраво виражене у земноводних i деяких органiзмiв з рiзко телоцитальною будовою яiць. В результатi у зародкiв таких тварин бластопор вiдсутнiй i архентерон не формуiться. Тiльки далi, коли макрозаходи дiлячись стають меншого розмiру, утворюiться порожнина -формируется зачаток первинного кишковика.

Мал. 5. Епiболiя (схема)
У чистому виглядi вказанi способи гаструляцii зустрiчаються украй рiдко. У кожному конкретному випадку ембрiогенезу, як правило, поiднуються декiлька типiв рухiв. В результатi прийнято видiляти останнiй, змiшаний, спосiб гаструляцii.
2. Провiзорнi органи зародка у птахiв
Провiзорнi, або тимчасовi, органи утворюються в ембрiогенезi ряду представникiв хребетних для забезпечення життiво важливих функцiй, таких, як дихання, живлення, видiлення, рух та iн. Недорозвиненi органи самого зародка ще не здатнi функцiонувати за призначенням, хоча обов'язково грають якусь роль в системi цiлiсного органiзму, що розвиваiться. Як тiльки зародок досягаi необхiдноi мiри зрiлостi, коли бiльшiсть органiв здатнi виконувати життiво важливi функцii, тимчасовi органи розсмоктуються або вiдкидаються.
Час утворення провiзорних органiв залежить вiд того, якi запаси поживних речовин були накопиченi в яйцеклiтинi i в яких умовах середовища вiдбуваiться розвиток зародка. У безхвостих земноводних, наприклад, завдяки достатнiй кiлькостi жовтка в яйцеклiтинi i тому, що розвиток йде у водi, зародок здiйснюi газообмiн i видiляi продукти дисимiляцii безпосередньо через оболонки яйця i досягаi стадii пуголовка. На цiй стадii утворюються провiзорнi органи дихання (зябра), травлення i рухи, пристосованi до водного способу життя. Перерахованi личинковi органи дають можливiсть пуголовку продовжити розвиток. Пiсля досягнення стану морфофункциональной зрiлостi органiв дорослого типу тимчасовi органи зникають в процесi метаморфоза.
У плазунiв i птахiв запасiв жовтка в яйцеклiтинi бiльше, але розвиток йде не у водi, а на сушi. У зв'язку з цим дуже рано виникаi потреба в забезпеченнi дихання i видiлення, а також в захистi вiд висихання. У них вже в ранньому ембрiогенезi, майже паралельно з нейруляцiiю, починаiться формування провiзорних органiв, таких, як амнiон, хорiон i жовтковий мiшок. Трохи пiзнiше формуiться аллантоис. У ссавцiв плацентарних цi ж провiзорнi органи утворюються ще ранiше, оскiльки в яйцеклiтинi дуже мало жовтка. Розвиток таких тварин вiдбуваiться внутрiутробний, утворення провiзорних органiв у них спiвпадаi за часом з перiодом гаструляцii.
Наявнiсть або вiдсутнiсть амнiона i iнших провiзорних органiв лежить в основi дiлення хребетних на двi групи: Amniota i Anamnia. Еволюцiйно бiльш древнi хребетнi, такi, що розвиваються виключно у водному середовищi i представленi такими класами, як Круглоротые, Риби i Земноводнi, не потребують додаткових водних i iнших оболонках зародка i складають групу анамний. До групи амниот вiдносять первичноназемных хребетних, тобто тих, у кого ембрiональний розвиток протiкаi в наземних умовах.
Це три класи: Плазуни, Птахи i Ссавцi. Вони i вищими хребетними, оскiльки мають скоординированные i високоефективнi системи органiв, що забезпечують iм iснування в найбiльш складних умовах, якими i умови сушi. Цi класи налiчують велику кiлькiсть видiв, що повторно перейшли у водне середовище. Таким чином, вищi хребетнi виявилися в змозi освоiти усi мiсця iснування. Подiбна досконалiсть була б неможливою, у тому числi i без внутрiшнього заплiднення i спецiальних провiзорних ембрiональних органiв.
У будовi i функцiях провiзорних органiв рiзних амниот багато спiльного. Характеризуючи в найзагальнiшому виглядi провiзорнi органи зародкiв вищих хребетних, званi також зародковими оболонками, слiд зазначити, що усi вони розвиваються з клiтинного матерiалу зародкових листкiв, що вже сформувалися. Деякi особливостi i в розвитку зародкових оболонок ссавцiв плацентарних, про що буде сказано нижче.
Амнiон i ектодермальним мiшком, що укладаi зародок i заповнений амниотической рiдиною. Амниотическая оболонка спецiалiзована для секрецii i поглинання амниотической рiдини, що омиваi зародок. Амнiон граi первинну роль в захистi зародка вiд висихання i вiд механiчних ушкоджень, створюючи для нього найбiльш сприятливе i природне водне середовище. Амнiон маi i мезодермальный шар з позазародковоi соматоплеври, який даi початок гладким м'язовим волокнам. Скорочення цих м'язiв викликають пульсацiю амнiона, а повiльнi коливальнi рухи, що повiдомляються при цьому зародку, мабуть, сприяють тому, що його зростаючi частини не заважають один одному.
Хорiон (сероза) - сама зовнiшня зародкова оболонка, прилегла до шкаралупи або материнських тканин, виникаюча, як i амнiон, з ектодерми i соматоплеври. Хорiон служить для обмiну мiж зародком i довкiллям. У яйцеродних видiв основна його функцiя - дихальний газообмiн; у ссавцiв вiн виконуi набагато бiльшi функцii, беручи участь окрiм дихання в живленнi, видiленнi, фiльтрацii i синтезi речовин, наприклад гормонiв.
Жовтковий мiшок маi энтодермальное походження, покритий вiсцелярною мезодермою i безпосередньо пов'язаний з кишковою трубкою зародка. У зародкiв з великою кiлькiстю жовтка вiн бере участь в живленнi. У птахiв, наприклад в спланхноплевре жовткового мiшка, розвиваiться судинна мережа. Жовток не проходить через жовткову протоку, що сполучаi мiшок з кишкою. Спочатку вiн переводиться в розчинну форму пiд дiiю травних ферментiв, що продукуються энтодермальными клiтинами стiнки мiшка. Потiм потрапляi в посудини i з кров'ю розноситься по усьому тiлу зародка.
Доля жовткового мiшка у рiзних тварин дещо рiзна. У птахiв до кiнця перiоду iнкубацii залишки жовткового мiшка вже знаходяться усерединi зародка, пiсля чого вiн швидко зникаi i до кiнця 6-оi доби пiсля вилуплення повнiстю розсмоктуiться.
Аллантоис розвиваiться дещо пiзнiше за iншi позазародковi органи. Вiн i мiшковидним вирощуванням вентральноi стiнки задньоi кишки. Отже, вiн утворений ентодермою зсередини i спланхноплеврой зовнi. У рептилiй i птахiв аллантоис швидко доростаi до хорiону i виконуi декiлька функцiй. Передусiм це вмiстище для сечовини i сечовоi кислоти, якi i кiнцевими продуктами обмiну азотвмiсних органiчних речовин. У аллантоисе добре розвинена судинна мережа, завдяки чому разом з хорiоном вiн бере участь в газообмiнi. При вилупленнi зовнiшня частина аллантоиса вiдкидаiться, а внутрiшня - зберiгаiться у виглядi сечового мiхура.
Доступнiше i простiше вивчити утворення i будову зародкових оболонок на прикладi зародка курки. На стадii нейрули три зародковi листки безпосередньо переходять вiд зародка до позазародковоi частини, нiяк не вiдмежовуючись. У мiру того як зародок набуваi форми, навколо нього утворюiться декiлька складок, якi як би пiдсiкають зародок, вiддiляють його вiд жовтка i встановлюють чiткi межi мiж зародком i позазародковими областями. Вони називаються тулубовими складками. (Мал.1).
Спочатку утворюiться головна складка. Вона пiдсiкаi знизу головну частину. Заднi кiнцi цiii складки переходжуватимуть в бiчнi тулубовi складки, що вiдмежовують тулуб зародка з бокiв. Хвостова складка вiдмежовуi заднiй кiнець зародка. Поступово звужуiться нiжка, що сполучаi середню кишку i жовтковий мiшок, утворюiться переднiй i заднiй вiддiли кишки.

Мал. 1 Утворення тулубових складок i зародкових оболонок у зародка курчати. А -продольный зрiз; Би -поперечный зрiз:
1 -эктодерма, 2 -мезодерма, 3 -зачаток мозку, 4 -глоточная мембрана, 5 -нервная трубка, 6-хорда, 7-клоачна мембрана, 8-хорион, 9-амнион, 10-экзоделом, 11-аллаитоис, 12-область пупка, 13-зачаток серця, 14-энтодерма, 15-закладка кишковика, 16-тулубовi складки, 17-жовтковий мiшок
Одночасно з ектодерми i прилеглоi до неi соматоплеври утворюiться спочатку головна складка (мал.), яка, як капюшон, наростаi на зародок згори. Кiнцi головноi складки утворюють з бокiв амниотические валики. Вони ростуть поверх зародка один назустрiч одному i зростаються, утворюючи вiдразу стiнки амнiона, прилеглого до зародка, i хорiону, що лежить зовнi.
Пiзнiше утворюiться аллантоис (мал.2). Загальний вигляд курячого зародка на 6-у добу iнкубацii зображений на мал.2. У рiзних ссавцiв процеси утворення провiзорних органiв бiльше або менше схожi на описаних вище у птахiв.

Мал. 2 Курячий зародок близько 40 ч iнкубацii: 1-головна складка амнiона, 2-нервова трубка, 3 - сомiти
Список лiтератури
1. Грин Н., Стаут У., Тейлор Д. Бiологiя / Н. Грин, У. Стаут, Д. Тейлор, пiд ред. Р. Сопера. - М.: Свiт, 2007. - Т. 1. - 368 с.
2. Грин Н., Стаут У., Тейлор Д. Бiологiя / Н. Грин, У. Стаут, Д. Тейлор, пiд ред. Р. Сопера. - М.: Свiт, 2009. - Т. 2. - 325 с.
3. М.А. Андрейчин, В.С. Копча Епiдемiологiя Терн. "Укр.медкнига"-2000р. стор. 349-356.
4. MED/96217734. Jacobsen H, Hanggi M, Ott M, Duncan IB, Owen S, Andreoni M, Vella S, Mous J. In vivo resistance to a human immunodeficiency virus type 1 proteinase inhibitor: mutations, kinetics, and frequencies. J Infect Dis. 1996 Jun;173(6):1379-87.
Вместе с этим смотрят:
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Бiологiчне рiзноманiття людських рас