Рецепторы как главное звено в деятельности сенсорной системы организма
Все живые организмы нуждаются в информации об окружающей среде как для поисков пищи и особей другого пола, так и при избегании разного рода опасностей. Кроме того, они должны ориентироваться в пространстве и оценивать его важнейшие свойства. Эту возможность обеспечивают сенсорные системы.
Разнообразные раздражители классифицируют, прежде всего, по модальности, т. е. по той форме энергии, которая свойственна каждому из них. Так, раздражители делят на механические, химические, тепловые, осмотические, световые, электрические и др. Эти раздражители передаются с помощью различных форм энергии; например, свет тАФ фотонами, химические раздражители тАФ молекулами и ионами, тепловые тАФ с помощью температуры, механические тАФ посредством механической формы энергии.
Кроме того, все раздражители независимо от их модальности подразделяются на адекватные и неадекватные. Адекватность раздражителя проявляется в том, что его пороговая интенсивность значительно ниже по сравнению с неадекватными раздражителями, например воздействие светового и механического стимулов на рецепторы глаза. Ощущение света возникает у человека, когда минимальная интенсивность светового раздражителя составляет всего 10 тАФ 10 Вт. Но ощущение вспышки можно вызвать и при механическом воздействии на глаз (это явление называют механическим фосфеном). Большинству людей оно знакомо по собственному опыту. Так, механическое давление на глазное яблоко воспринимается как свет, а от резкого удара по глазам Вл из глаз сыплются искрыВ», хотя никакого света при этом не было. Для возникновения вспышки света механическим путем мощность стимула должна быть более 10~ Вт.
Следовательно, разница между световым и механическим пороговым раздражителями для рецепторов глаза человека достигает 13тАФ14 порядков.
Деятельность любой сенсорной системы начинается с восприятия рецепторами внешней для мозга физической и химической энергии, трансформации ее в нервные импульсы и передачи их в мозг через цепи нейронов, образующих ряд уровней.
Данная работа посвящена рецепторам и рассматривает основные механизмы физиологии рецепции, которые свойственны всем сенсорным модальностям.
рецептор адаптация сенсор система
1. Рецепторы и их классификация
Рецепторы представляют собой конечные специализированные образования, предназначенные для трансформации энергии различных видов раздражителей в специфическую активность нервной системы.
Рецепторные клетки отличаются от остальных, по крайней мере, в двух отношениях. Во-первых, энергия раздражителя служит для них лишь стимулом к запуску процессов, совершаемых за счет потенциальной энергии, которая накоплена вследствие обменных реакций в самой клетке. Во-вторых, рецепторная клетка обладает на выходе электрической энергией, обязательно передаваемой другим клеткам, которые сами не способны воспринимать энергию данного внешнего воздействия.
Основной структурной единицей большинства рецепторных аппаратов является клетка, снабженная подвижными волосками, или ресничками. Эти волоски представляют собой как бы периферические подвижные антенны, действующие подобно усилителям по отношению к воспринимаемым раздражителям и участвующие в трансформации раздражителя в нервную сигнализацию. Волоски содержат в своем составе 9 пар периферических и 2 центральные фибриллы. Центральные фибриллы выполняют опорную роль, а периферические, содержащие миозиноподобные макромолекулы, сокращаются под воздействием АТФ. Благодаря их автоматическим движениям осуществляются непрерывные поиски адекватного стимула и обеспечиваются наилучшие условия для взаимодействия с ним. Следовательно, в одной и той же клетке представлены и собственно рецепторная, и моторная функции.
Другая сторона деятельности рецепторных элементов заключается во взаимодействии энергии внешнего стимула с поверхностью антенн, которые покрыты мембраной (мембрана образована из двойного слоя липидов, ограниченного с обеих сторон слоем белковых молекул). Специфической особенностью рецепторных мембран является включение в их состав биологически активных веществ тАФ пигментов, ферментов, ацетилхолинэстеразы и др.
Следовательно, общий механизм рецепции слагается из механо-химических молекулярных процессов, обеспечивающих движение антенн, и общих биохимических циклов при взаимодействии специфического стимула с рецепторными мембранами антенн.
Однако не следует думать, что этой схемой ограничивается восприятие стимула рецепторной клеткой, У некоторых рецепторов во взаимодействии со стимулом принимает участие вся клетка (например, хеморецелторные клетки, чувствительные к напряжению кислорода в крови), у других (вкусовые луковицы позвоночных), восприятие осуществляется микроворсинками. В большей части рецепторов кожи, внутренних органов и мышц участки преобразования стимула находятся в окончаниях нервных волокон.
Пороги восприятия высокоспециализированными рецепторами адекватных стимулов чрезвычайно низки. Для возбуждения фоторецептора достаточно одного кванта света, обонятельные рецепторы информируют организм о появлении в атмосфере единичных молекул пахучих веществ, волосковые рецепторы лабиринта способны обнаружить движение примерно такое же малое, как диаметр атома водорода. По характеру взаимодействия раздражителей всю совокупность рецепторов подразделяют на экстероцепторы, воспринимающие раздражения внешних агентов, и интероцепторы, сигнализирующие о раздражителях внутренней среды. К первым относят такие высокоспециализированные образования, как рецепторы органов слуха, зрения, обоняния, вкуса, осязания; ко вторым тАФ рецепторы внутренних органов. Одной из разновидностей интероцепторов следует считать проприоцепторы (рецепторы опорно-двигательного аппарата).
У экстероцспторов в большей степени выражена так называемая специализация, под которой понимают высокую избирательную чувствительность к адекватному раздражителю (закон специфической перепой энергии Мюллера). Обладая чрезвычайно высокой чувствительностью к адекватному раздражителю, экстероцепторы, как правило, могут реагировать и на неадекватные стимулы, но лишь на очень интенсивные. Поэтому принято считать экстероцепторы мономодальными рецепторными приборами.
Среди интероцепторов также есть мономодальные образования, например хеморецепторы каротидной зоны, предназначенные для химического анализа крови, направляющейся к мозгу. Однако большинство интероцепторов являются полимодальными, т. е. способными реагировать не на один, а на несколько разных по модальности раздражителей, например на температурные, химические и механические. Разница в порогах восприятия адекватных и неадекватных раздражителей у полимодальных рецепторов не столь ярко выражена, как у мономодальных.
Наиболее понятная и удобная классификация рецепторов исходит из различной модальности воспринимаемых ими раздражителей. В соответствии с этим разнообразием все рецепторы живых организмов можно разбить на несколько групп.
1. Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Восприятие механического раздражителя необходимо как самым низшим организмам, таким, как бактерии и простейшие, так и высокоорганизованным позвоночным животным. У беспозвоночных они представлены первичной механочувствительностью всей поверхностной мембраны (бактерии, простейшие) и специализированными рецепторами, выполняющими экстеро- и про-приоцептивные функции у многоклеточных. У позвоночных механо-рецепторы подразделяются на рецепторы кожи, сердечно-сосудистой системы, внутренних органов, опорно-двигательного аппарата и акустико-латеральной системы. Механорецепторную функцию различных тканей и органов выполняют рецепторы ареснитчатого типа, тогда как в акустико-латеральной системе рецепторными клетками являются волосково-реснитчатые. Механорецепторы представляют периферические отделы соматической, скелетно-мышечной, слуховой и вестибулярной сенсорных систем, а также боковой линии.
2. Терморецепторы воспринимают температурные раздражения. Они объединяют рецепторы кожи и внутренних органов, а также центральные термочувствительные нейроны. У позвоночных терморецепторы подразделяются на холодовые и тепловые; они обнаруживают тепловое излучение косвенно по его влиянию на температуру кожи. У некоторых позвоночных (гремучие змеи) имеются специализированные рецепторы, непосредственно воспринимающие инфракрасные лучи.
3. Хеморецепторы чувствительны к действию химических агентов. У наземных животных они образуют периферические отделы обонятельной и вкусовой сенсорных систем, тогда как для водных животных эти понятия теряют смысл, что заставляет использовать термин хемо-рецепция или химическая чувствительность. Интероцепторы (сосудистые и тканевые) участвуют в оценке химического состава внутренней среды и связаны с работой висцерального анализатора.
4. Фоторецепторы воспринимают световую энергию. Они представлены цилиарными рецепторами, т.е. производными клетки со жгутиком, и рабдомерными, у которых жгутик отсутствует, а собственно фоторецепторная часть клетки образована совокупностью микровилл.
5. Электрорецепторы чувствительны к действию электромагнитных колебаний. Они обнаружены в составе боковой линии у круглоротых, пластиножзберных, многих костистых рыб и некоторых хвостатых амфибий. К ним относятся ампулированные и бугорковые электрочувствительные рецепторные органы.
6. Болевые (ноцицептивные) рецепторы воспринимают болевые раздражения. Однако наряду со специализированными нервными окончаниями болевые стимулы могут восприниматься также и другими типами сенсорных аппаратов.
На уровне молекул и клеточных мембран основные рецепторные механизмы в пределах данной модальности имеют много общих свойств у разных типов и видов животных. Однако в зависимости от образа жизни, среды обитания и ряда других биологических факторов рецепторные приборы организмов могут существенно различаться.
У некоторых животных организм может быть вообще лишен многих рецепторов, например в случае крайней адаптации ленточного червя к паразитическому существованию в кишечнике хозяина.
В других случаях живые организмы не могут воспринимать ту или иную модальность или имеют ограниченный диапазон для ее анализа.
Например, у человека не обнаружены электрорецепторы, существующие у рыб; нет рецепторов, воспринимающих прямое инфракрасное излучение, как у гремучей змеи; глаз человека не воспринимает поляризацию света, как глаза некоторых насекомых; его ухо не ощущает ультразвуковых колебаний, как слуховой аппарат летучих мышей и многих ночных млекопитающих.
Однако бесспорно, что рецепторные аппараты обеспечивают каждый организм достаточным количеством информации, которая необходима для его нормального существования, так как они приспособлены к восприятию именно тех сигналов, которые существенны именно для данного вида животного.
Важнейшее свойство рецепторов тАФ избирательная чувствительность к адекватным раздражителям. Выраженность этого свойства у тех или иных рецепторных аппаратов обусловлена в значительной мере их структурными особенностями. На основании этого все рецепторы могут быть разделены на две группы: первичные (первичночувствующие) и вторичные (вторичночувствующие) (рис. 1).
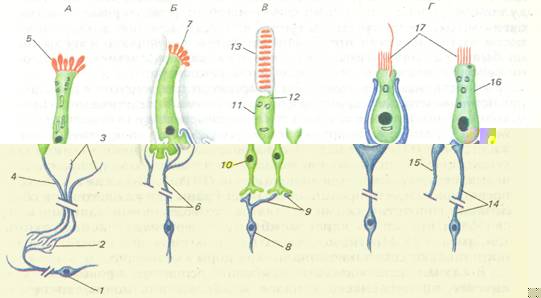
Рис. 1. Специализированные первичные (А) и вторичные (БтАФГ) рецепторные клетки позвоночных. А тАФ обонятельный рецептор; Б тАФ вкусовой; В тАФ фоторецептор; Г тАФ вестибулярный и слуховой:
1 тАФ митральная клетка, 2 тАФ обонятельный клубочек, 3 тАФ обонятельные нити, 4 тАФ аксон, 5 тАФ реснички, 6 тАФ волокна chorda tympani 7 тАФ микроворсинки, 8 тАФ биполярная клетка, 9 тАФ волокна, 10 тАФ ядро, 11 тАФ внутренний членик, 12 тАФ рудимент реснички, 13 тАФ наружный членик, 14 тАФ эфферентное нервное волокно. 15 тАФ эфферентный аксон, 16 тАФ наружная волосковая клетка (улитка), 17 тАФ волоски
К первичным относят такие рецепторные аппараты, у которых действие адекватного стимула осуществляется непосредственно периферическим отростком сенсорного нейрона, который, таким образом, первично встречается с раздражителем. Этот сенсорный нейрон находится на периферии, а не в центральной нервной системе, и представляет собой преобразованный в ходе эволюции биполярный нейрон, на одном полюсе которого расположен дендрит с ресничкой или дендритными отростками, а на другом тАФ центральный отросток тАФ аксон, по которому возбуждение передается в соответствующий центр. К вторичным рецепторам относят такие рецепторы, у которых между окончаниями сенсорного нейрона и точкой приложения стимула располагается дополнительная специализированная (рецепти-рующая) клетка ненервного происхождения. Возбуждение, возникающее в рецептирующей клетке, передается через синапс на сенсорный нейрон. Следовательно, сенсорный нейрон возбуждается уже не первично внешним стимулом, а опосредованно (вторично) благодаря воздействию рецептирующих клеток. Последние не имеют периферических и центральных отростков, но восприятие стимула у них осуществляется с помощью жгутикообразных волосков.
Первичные рецепторы, впервые появляющиеся уже у кишечнополостных, являются основным универсальным типом рецепторных элементов, с которыми связаны все виды рецепции у беспозвоночных. Интересно отметить, что вторичночувствующие эпителиальные клетки обнаружены недавно в органах гравитации у таких низкоорганизованных животных, как гребневики.
У позвоночных животных первичные рецепторы представлены тканевыми рецепторами и проприоцепторами, а также терморецепторами и обонятельными клетками. К вторичным рецепторам следует отнести рецепторные элементы органов боковой линии (механо- и электрорецепторы), волосковые клетки внутреннего уха, рецепторные клетки вкусовых луковиц и фоторецепторы глаза позвоночных.
2. Адаптация рецепторов
Постоянно действующий стимул лишь в редких случаях создает в рецепторах постоянный уровень возбуждения на неопределенный срок. Чаще при длительном раздражении возбуждение слабеет в большей или меньшей степени. Это явление называется адаптацией рецепторов. В его основе лежат весьма сложные процессы, протекающие в рецепторах и в центральных отделах нервной системы. Субъективно адаптация проявляется по отношению к воздействию постоянного раздражителя. Например, войдя в прокуренное помещение, человек через несколько минут перестает ощущать столь резкий вначале запах табака. Точно так же мы не замечаем непрерывного давления на кожу привычной одежды или яркого света, который вначале нас ослепляет.
В зависимости от способности изменять свою активность в ходе действия длительного раздражения все рецепторы могут быть разделены на две группы: быстро адаптирующиеся, или фазные, и медленно адаптирующиеся, или тонические (рис.3). Существует также и промежуточный тип рецепторов тАФ фазнотоничсские.
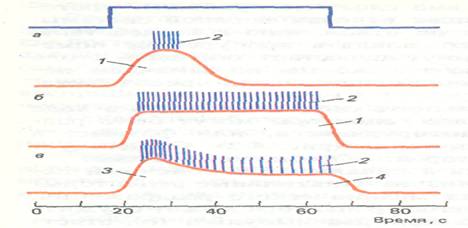
Рис. 2. Реакции фазных (а), тонических (б) и фазно-тонических (в) рецепторов при стационарном раздражении: I тАФ рецепторный потенциал, 2 тАФ спайковая активность, 3 тАФ фазовый компонент ряцепторного потенциала, 4 тАФ тонический компонент рецепторного потенциала; вверху показана отметка раздражения
Фазные рецепторы возбуждаются в начальный и конечный периоды деформации их мембран при низких (тактильные рецепторы) и высоких (фонорецепторы) частотах механического раздражения. Примером очень быстро адаптирующегося рецептора может служить тельце Пачини, генерирующее всего один-два импульса (on-ответ) в момент начала действия стационарной деформации и один-два ПД (оff-ответ) в момент выключения раздражения.
Тонические рецепторы возбуждаются в течение всего периода деформации их мембран. В начальный момент действия раздражителя наблюдаются высокочастотные разряды, после которых импульсация устанавливается на более низком уровне соответственно величине приложенного давления и сохраняется в течение всего времени действия раздражителя. Примером тонического рецептора является медленный рецептор растяжения у ракообразных, активность которого может сохраняться на одном уровне в течение нескольких часов.
В фазно-тонических рецепторах электрические процессы регистрируются все время, пока длится раздражение, однако амплитуда РП и частота ПД резко снижаются при длительном его воздействии. Рецепторный потенциал таких рецепторов имеет два четко выраженных компонента: начальный тАФ фазный, последующий тАФ тонический.
Таким образом, организм, имея в своем составе все три типа рецепторов, получает информацию о начальных и конечных моментах воздействия раздражителя, а также о его силе и длительности.
На основе деятельности тонических рецепторов может постоянно анализироваться и прослеживаться уровень сенсорного воздействия. Фазный тип реагирования обеспечивает возможность отражать моменты изменения этого уровня, а фазно-тонический тАФ объединяет в себе те и другие свойства.
Адаптационные процессы в рецепторах могут определяться внешними и внутренними факторами. В качестве внешнего фактора в механизме адаптации могут выступать свойства вспомогательных структур. Например, как показали специальные эксперименты, основной причиной быстрой адаптации телец Пачини являются свойства впомогательных структур (капсулы рецептора), которые не пропускают к нервному окончанию статической составляющей раздражающего действия. Динамическая же составляющая стимула хорошо проходит через элементы капсулы, хотя и уменьшается несколько по амплитуде. Однако после удаления капсулы рецептор начинает вести себя как тонический и генерировать РП в течение действия достаточно продолжительного стимула,
Внутренние факторы механизма адаптации связаны с изменениями физико-химических процессов в самом рецепторе. Сравнительный анализ характеристик мембранных ионных каналов быстро адаптирующихся и медленно адаптирующихся волокон свидетельствует о различии их натриевых и калиевых систем. Различный набор этих ионных каналов может, по-видимому, определять фазный или тонический тип активности сенсорного прибора.
В явлениях адаптации важную роль могут играть также эфферентные влияния от расположенных выше нервных центров. При наличии тормозной эфферентной регуляции процессы адаптации в рецепторах ускоряются. Например, стационарная импульсация, поступающая от центров к рецепторам растяжения ракообразных, способна превратить ответ тонического рецептора в фазный.
Таким образом, из представленного следует, что для адаптации рецепторов нет единого общего закона и в каждой сенсорной системе может быть свое сочетание факторов, которое и определяет изменение возбудительного процесса.
Заключение
Рецепторам принадлежит важнейшая роль в получении организмом информации о внешней и внутренней средах. Благодаря их большому разнообразию в организме животные и человек способны воспринимать стимулы разных модальностей. Процесс передачи сенсорного сообщения сопровождается многократным преобразованием и перекодированием и завершается общим анализом и синтезом (опознаванием образа). После этого происходит выбор или разработка программы ответной реакции- организма. Без информации, поступающей в мозг, не могут осуществляться простые и сложные рефлекторные акты вплоть до психической деятельности человека.
Литература
1. Батуев А.С. Куликов Г.А. Введение в физиологию сенсорных систем. тАУ М.: ПЕДАГОГИКА, 1983. тАУ 324 с.: ил.
2. Большая Советская Энциклопедия (В 30 томах). Т. 22 Ремень-Сафи /Гл. ред. А.М. Прохоров. тАУ М.: Советская Энциклопедия, 1975. тАУ 628 с.: ил
3. Кадель В.Л. Физиология органов чувств. тАУ М.: Высш. шк.,1975. тАУ 253 с.: ил
4. Общий курс физиологии человека и животных. В 2 кн. Кн. 1. Физиология нервной, мышечной и сенсорной систем: Учеб. для биол. и медиц. спец. вузов /Под ред А.Д. Ноздрачева. тАУ М.: Высш. шк., 1991. тАУ 512 с.: ил.
5. Энциклопедический словарь юного биолога/Сост. М.Е. Аспиз. тАУ М.: Педагогика, 1986. тАУ 352 с.: ил.
Вместе с этим смотрят:
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Бiологiчне рiзноманiття людських рас
Бiологiя iндивiдуального розвитку
Бiологiя лева
Банан