Участие прокариот в круговороте серы
Прокариоты (бактерии и археи) имеют исключительное значение для жизни на Земле тАУ им принадлежит основополагающая роль в циклических превращениях основных элементов, необходимых для жизни (углерод, кислород, азот, сера, фосфор). Циклические превращения элементов, из которых построены живые организмы, в совокупности представляют круговорот веществ. В настоящее время неоспоримо доказано, что определенные этапы циклов осуществляют исключительно прокариоты, обеспечивая замкнутость циклов основных биогенных элементов в биосфере. По В.И. Вернадскому (один из основоположников учения о биосфере), Влбиосфера представляет оболочку жизни тАУ область существования живого веществаВ».
1. Круговорот серы
Сера тАУ биогенный элемент, необходимый компонент живой материи. Она содержится в белках в составе аминокислот, содержание серы в белках составляет 0,8тАУ2,4%. Сера также входит в состав витаминов, гликозидов, коферментов, имеется в растительных эфирных маслах. Сера в изобилии присутствует в земной коре, в углях, сланцах, нефтях, природных газах.
Сера относится к элементам с переменной валентностью. Это обеспечивает ее подвижность. В виде неорганических соединений сера бывает в окисленной форме (сульфаты, политионаты), восстановленной форме (сульфиды) и молекулярной, осуществляя активный окислительно-восстановительный цикл. В природе сера претерпевает разнообразные химические и биологические превращения, переходя из неорганических соединений в органические и обратно, меняя валентность в пределах от тАУ 2 до +6.
Циклические превращения соединений серы называются круговоротом серы.
Цикл превращения серы включает окислительные и восстановительные звенья, а также превращения серы без изменения ее валентности. Окислительная часть круговорота серы включает стадии, которые могут протекать в зависимости от условий как чисто химически, так и с участием организмов, главным образом микроорганизмов (это организмы, не видимые невооруженным глазом, включающие микроскопические эукариоты: грибы, водоросли, простейшие и все прокариоты). Восстановительная часть круговорота серы осуществляется преимущественно биологическим путем с доминирующей ролью прокариот в этом процессе, при этом осуществляется восстановление атома серы из состояния максимального окисления (+ 6) до максимального восстановления (тАУ2). Однако не всегда этот процесс происходит до конца, и в среде нередко обнаруживаются не полностью окисленные продукты: элементная сера, политионаты, сульфит.
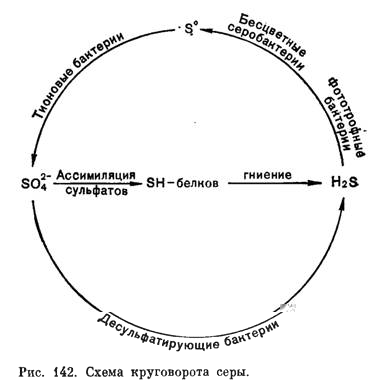
Таким образом, цикл серы, так же как и круговорот веществ, невозможен без участия прокариот, обеспечивающих замкнутость цикла.
2. Восстановительная ветвь
Ассимиляция сульфата.
Сульфат используется в качестве источника серы почти всеми растениями и микроорганизмами. Сульфат при ассимиляции восстанавливается, чтобы сера могла включиться в органические соединения, так как в живых организмах сера встречается почти исключительно в восстановленной форме в виде сульфгидрильных (-SH) или дисульфидных (-S-S-) групп. В обоих случаях ассимилируется ровно столько питательных веществ, содержащих серу, сколько их необходимо для роста организма, поэтому никакие восстановленные продукты метаболизма серы не выделяются в окружающую среду. В результате биосинтеза сера включается в основном в состав серосодержащих аминокислот: цистин, цистеин, метионин. Вовлечение сульфатов в состав серосодержащих органических веществ носит название ассимиляционной сульфатредукции.
Превращение органических соединений серы с образованием H2S.
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
При минерализации органических серосодержащих соединений сера освобождается в неорганической восстановленной форме в виде H2S. В освобождении серы из органических серосодержащих соединений (продукты метаболизма живых существ, отмершие растительные и животные остатки) принимают участие сапрофитные микроорганизмы, способные к аммонификации. При аммонификации серосодержащие белки и нуклеиновые кислоты разлагаются с образованием СО2, мочевины, органических кислот, аминов и, что важно для цикла серы, H2S и меркаптанов (тиоспирты). Меркаптаны в аэробных условиях также окисляются с выделением H2S.
Разрушение белков микроорганизмами начинается как внеклеточный процесс. При этом белки гидролизуются протолитическими экзоферментами до более мелких молекул, способных проникать внутрь клетки и расщепляться внутриклеточными протеазами до аминокислот, которые могут подвергаться дальнейшему расщеплению.
Прямое образование H2S из сульфата и элементной серы.
Процессы образования в биосфере сероводорода связывают в основном с деятельностью сульфатредуцирующих бактерий, имеющих большое значение для глобального круговорота серы. Сульфатредуцирующие бактерии осуществляют диссимиляционную сульфатредукцию, представляющую собой анаэробное дыхание, при котором сульфат служит конечным акцептором электронов (вместо кислорода) при окислении органических веществ или молекулярного водорода. Поэтому энергетический тип обмена у сульфатредуцирующих бактерий часто называют сульфатным дыханием. Схематически процесс восстановления сульфатов при диссимиляционной сульфатредукции можно представить следующим образом: SO42-→SO32-→S3O62-→S2O32-→S2-.
Ферментативная система, участвующая в восстановлении сульфата, состоит из двух частей: первая восстанавливает сульфат в сульфит в АТФ-зависимом процессе, а вторая восстанавливает сульфит в сульфид путем шестиэлектронного переноса. Именно последняя реакция, являясь диссимиляторной, снабжает клетку энергией.
Сульфатредуцирующие бактерии преимущественно облигатные анаэробные бактерии. Геохимическая роль сульфатредуцирующих бактерий чрезвычайно велика, поскольку благодаря их деятельности инертное соединение тАУ сульфат в анаэробной зоне в больших масштабах вовлекается в биологический круговорот серы.
Деятельность сульфатредуцирующих бактерий особенно заметна в иле на дне прудов и ручьев, в болотах и вдоль побережья моря. Так как концентрация сульфата в морской воде относительно высока, восстановление сульфата тАУ важный фактор минерализации органического вещества на морских отмелях. Признаками такой минерализации служит запах H2S и черный как смоль ил, в котором протекает этот процесс. Черный цвет ила обусловлен присутствием в нем больших количеств сульфида двухвалентного железа. Некоторые береговые области, где накопление органического вещества ведет к особенно интенсивному восстановлению сульфата, практически безжизненны из-за токсического действия H2S.
Сульфатредуцирующие бактерии тАУ это физиологическая, а не систематическая группа, так как к ним относятся бактерии из разных таксономических групп, способные осуществлять один физиологический процесс тАУ анаэробное дыхание в присутствии сульфатов, например бактерии родов Desulfovibrio (вибрионы), Desulfotomaculum (спорообразующие палочки). Среди сульфатредуцирующих организмов обнаружены и археи. Сульфатредуцирующие бактерии могут осуществлять рост за счет не только восстановления сульфатов, но и тиосульфата, сульфита, элементной серы и других соединений серы.
Для некоторых сульфатредуцирующих бактерий был показан преимущественно новый тип метаболизма серы. Эти бактерии способны получать энергию при росте на органических субстратах не только за счет сульфатредукции, но и в результате диспропорционирования тиосульфата, сульфита, дитионита с образованием сульфата и сульфида.
Суммарная реакция диспропорционирования тиосульфата:
S2O32- + H20 → SO42- + HS- + H+
Go = тАУ 21,9 кДж/моль S2O32-
Суммарная реакция диспропорционирования сульфита:
4SO32- + H+ → 3SO42- + HS-
Go = тАУ 58,9 кДж/ моль SO32-
Обнаружено, что бактерии, осуществляющие диспропорционирование тиосульфата, широко распространены в морских осадках.
Сероводород может образовываться также при восстановлении элементной серы. На сегодня известны два механизма образования сероводорода из молекулярной серы. В первом случае бактерии и археи используют молекулярную серу как акцептор электронов при анаэробном дыхании (диссимиляционная сероредукция), в ходе которого синтезируется АТФ. Диссимиляционная сероредукция тАУ это ферментативный процесс, который осуществляют как мезофильные, так и термофильные прокариоты. Во втором случае микроорганизмы (дрожжи и прокариоты) используют серу лишь для сброса электронов, освобождающихся при брожении (облегченное брожение). Это футильный (холостой) сброс электронов, который не сопровождается синтезом АТФ.
Большое значение восстановление молекулярной серы в сероводород имеет именно для термофильных микроорганизмов, обитающих в гидротермах, где элементная сера является одной из наиболее значимых форм серы и где соединения серы имеют исходно вулканическое происхождение. Биоценозы гидротерм представляют собой уникальные сообщества живых существ. Развиваясь при высоких температурах (от 45тАУ50 до 100В°С), они образуются в основном прокариотами тАУ бактериями и археями. Подавляющее большинство микроорганизмов, входящих в состав микробных сообществ гидротерм, не встречаются в других местах. Микробные сообщества гидротерм относят к наиболее древним биоценозам Земли.
3. Окислительная ветвь
Эта часть серного цикла может состоять как целиком из реакций неорганических соединений серы: S2- → nS2- → S0 → S2O32- → SO32- → SO42-, так и включать реакции органических форм. Атом серы органических сульфидов обычно окисляется после отделения в виде S2 тАУ по неорганическому пути, хотя возможен и чисто органический путь окисления, когда атом серы окисляется, находясь в составе органических соединений, например цистеин → цистин. Большинство реакций окисления серных соединений может протекать без участия микроорганизмов в присутствии сильных окислителей (H2O2, O3, кислородные радикалы), однако микробное окисление гораздо более эффективно, особенно при низких концентрациях реагента.
Среди микроорганизмов, способных к окислению серных соединений, выделяются три основные физиологические группы: анаэробные фототрофные бактерии, аэробные и факультативно анаэробные литотрофные бактерии, археи и различные гетеротрофные микроорганизмы.
Анаэробные фототрофные бактерии.
Это специфическая группа бактерий, осуществляющая анаэробный (аноксигенный) фотосинтез, использующая в качестве доноров электронов различные восстановленные серные соединения вместо H2O, как это происходит при аэробном (оксигенном) фотосинтезе у растений, цианобактерий. Наиболее часто продуктами окисления восстановленных соединений серы являются SO42- и S0, последняя может накапливаться внутриклеточно у некоторых фототрофных бактерий.
В систематическом отношении фототрофные бактерии делятся на несколько групп: серные и несерные пурпурные и зеленые бактерии, гелиобактерии, эритробактерии (рис. 123). Фототрофные бактерии пигментированы и могут быть окрашены в коричневый, зеленый, пурпурный цвета. Фототрофные бактерии, осуществляющие аноксигенный фотосинтез, содержат бактериохлорофилл и каротиноиды, которые и придают им соответствующую окраску. Данные бактерии доминируют в окислении сульфида в анаэробной водной толще неглубоких водоемов, таких, как озера, пруды, лагуны, минеральные источники, куда проникает достаточно света. Исключение оставляют эритробактерии, которые являются облигатными аэробами и осуществляют аноксигенный тип фотосинтеза в аэробных условиях.
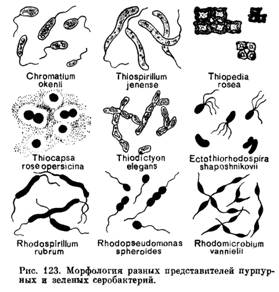
Литотрофные сероокисляющие бактерии и археи. Серобактерии и тионовые бактерии.
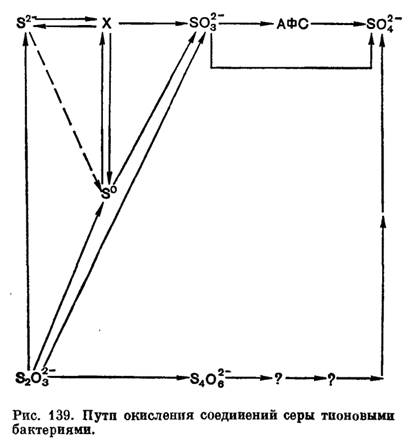
Некоторые прокариоты в присутствии восстановленных соединений серы способны к литотрофному росту. При литотрофном росте восстановленные соединения серы или другие восстановленные неорганические соединения (NH3, H2, CO, Fe2+) являются донорами электронов в энергетическом метаболизме, то есть при их окислении в электрон-транспортной цепи синтезируется АТФ. К литотрофному росту способны только прокариоты. У литотрофных серозависимых прокариот реакции окисления восстановленных соединений серы служат источником энергии. Окислительные реакции осуществляются при участии специфических ферментных систем. Окисление литотрофными прокариотами разных соединений серы и самой серы обычно ведет к образованию сульфатов. Однако не всегда этот процесс происходит до конца, и в среде нередко обнаруживаются не полностью окисленные продукты (S0, SO32-, S2O32-, S4O62-). Группа литотрофных серозависимых прокариот в основном включает представителей тионовых бактерий, экстремально термоацидофильных серозависимых архей, водородных бактерий, бесцветных серобактерий, а также некоторых представителей пурпурных бактерий, способных к литотрофному росту за счет окисления серных соединений в темноте.
Давно известно, что в сероводородных источниках и других водоемах, содержащих сероводород, как правило, встречаются в большом количестве неокрашенные микроорганизмы, в клетках которых обнаруживаются капли серы. В местах, где концентрация сероводорода сравнительно невелика (меньше 50 мг/л), такие микроорганизмы, получившие название бесцветных серобактерий, часто образуют массовые скопления в виде пленок, белых налетов и других обрастаний. С.Н. Виноградский (1887) доказал, что сера, откладываемая в клетках одного из типичных представителей серобактерий, а именно Beggiatoa, образуется из сероводорода и может окисляться этим микроорганизмом до серной кислоты. В систематическом отношении бесцветные серобактерии, видимо, гетерогенная группа и далеко не все из описанных видов и даже родов можно считать твердо установленными. По морфологии, характеру движения, способу размножения и строению клеток ряд представителей бесцветных серобактерий, как многоклеточные, так и одноклеточные (Beggiatoa, Thiothrix, Thiospirillopsis, Thioploca, Achromatium) проявляют большое сходство с синезелеными водорослями. Некоторые исследователи, в частности Прингсхейм (Pringsheim, 1963), рассматривают эти микроорганизмы как бесцветные их варианты. Аналогом Beggiatoa считают сине-зеленую водоросль Oscillatoria, Thiothrix тАУ Rivularia, Thiospirillopsis тАУ Spirulina, a Achromatium похож на Synechococcus. Поскольку сине-зеленые водоросли сейчас причисляют к бактериям, то их сближение с бесцветными серобактериями становится все более обоснованным. Следует также отметить, что у некоторых сине-зеленых водорослей обнаружена способность откладывать в клетках серу, хотя один этот признак мало что дает для систематики микроорганизмов. Микроорганизмы, относящиеся к бесцветным серобактериям, встречаются и в пресных и в соленых водоемах. Некоторые из них хорошо растут при низкой температуре, другие (Thiospirillum pistiense) развиваются в термальных серных источниках при температуре более 50В°С. Подвижные формы обладают хемотаксисом и могут перемещаться в места с оптимальным содержанием кислорода и сероводорода.
Большинство так называемых тионовых бактерий тАУ типичные хемоавтотрофы, т.е. они используют восстановленные соединения серы не только как Н-доноры, но и в качестве источников энергии и способны расти на чисто минеральных средах, ассимилируя углекислоту. Такие бактерии были впервые выделены из воды Неаполитанского залива (Натансон, 1902) и получили название Thiobacillus (Бейеринк, 1904). К настоящему времени описано много видов автотрофных тиобацилл, выделенных из разных водоемов, почвы, а также из месторождений серы и разных металлов.
На основании некоторых, главным образом физиологических, особенностей (способность окислять различные соединения серы и другие неорганические субстраты, использовать органические соединения, расти в зависимости от значений рН среды, отношение к кислороду и др.) выделяют значительное число видов тиобацилл. Из них считаются наиболее твердо установленными следующие: Thiobacillus thioparus, Т. thiooxidans, Т. denitrificans, Т. thiocyanoxidans, Т. neapolitanus, Т. intermedius, Т. novellus, Т. ferrooxidanas. Все эти микроорганизмы растут на простых минеральных средах, куда часто вносят бикарбонат. Источником азота обычно служат соли аммония. Некоторые виды и штаммы способны ассимилировать как источники азота нитраты. Показана также способность отдельных представителей использовать мочевину и аминокислоты. Штаммы, выделенные из соленых водоемов, требуют для роста хлористого патрия, причем известны галофилы, способные расти в насыщенном растворе NaCl. Оптимальная температура для роста большинства выделенных микроорганизмов рода Thiobacillus около 30В°С. Однако в природных условиях они встречаются в термальных источниках при температуре до 55В°С. Недавно выделена чистая культура Thiobacillus sp. с оптимумом для роста при 50В°С.
По отношению к кислотности среды тиобациллы подразделяются на организмы, хорошо растущие при нейтральной или даже щелочной реакции, и виды, являющиеся ацидофильными, т.е. развивающиеся в кислой среде и выдерживающие очень низкое значение рН.
К первой группе относятся такие виды, как: Т. thioparus, Т. denitrificans, Т. novellus, Т. thiocyanoxidans, Т. neapolitanus. Для этих микроорганизмов оптимальное значение рН приходится на область 6,0тАУ9,0, а зона значений рН, при которых возможен их рост, тАУ от 3,0тАУ6,0 до 10,0тАУ11,0, причем для разных видов и штаммов оптимальные значения рН и. область активной кислотности, в которой наблюдается их рост, могут заметно различаться.
Ко второй группе принадлежат Т. thiooxidans, Т. ferroxidans, Т. intermedius. Для этих микроорганизмов оптимальное значение рН 2,0тАУ4,0, а рост возможен при рН от 0,5тАУ2,0 до 5,0тАУ7,0. Наиболее ацидофильными организмами являются два первых вида. Обе эти бактерии растут при значениях рН не более 5,0. В то же время показано, что Т. thiooxidans сохраняет жизнеспособность при значении рН, близком к 0, что соответствует 1,0 н. раствору серной кислоты. Это, пожалуй, самый ацидофильный микроорганизм, который известен исследователям.
Большинство тионовых бактерий растет только в присутствии кислорода, хотя рост некоторых представителей возможен при низком его содержании. Но известны и факультативные анаэробы. К ним относится Т. denitrificans. В аэробных условиях эти бактерии ведут окислительные процессы с участием молекулярного кислорода, в анаэробных переключаются на денитрификацию и восстанавливают нитраты до молекулярного азота. Интересно отметить, что ассимилировать нитраты как источники азота Т. denitrificans, подобно Paracoccus denitrificans, не может и требует для роста в качестве источника азота аммония.
Тионовые бактерии способны окислять такие соединения серы, как сероводород, сульфиды, сульфит, тиосульфат, тетратионат, тиоцианаг (роданид), дитионит, а также молекулярную серу, с образованием при полном их окисления сульфатов. Однако способности отдельных видов не вполне одинаковы. Кроме того, не всегда легко установить, какие именно соединения серы окисляются биологическим путем, так как многие из них неустойчивы при низком значении рН и могут также окисляться кислородом воздуха.
Все виды, растущие при нейтральной и щелочной реакции среды, как правило, окисляют сероводород, серу и тиосульфат. Характерным признаком Т. thiocyanoxidans является способность окислять роданид. На этом основании его выделяют в отдельный вид, так как по другим признакам этот микроорганизм похож на Т. thioparus. Такую ацидофильную бактерию, как Т. thiooxidans, культивируют обычно на среде, содержащей молекулярную серу. Вопрос о способности данного вида окислять сероводород и другие соединения серы окончательно не решен, так как в кислых условиях эти соединения неустойчивы. В отношении Т. ferroxidans есть данные, что эти бактерии могут окислять как молекулярную серу, так и различные ее соединения, а именно: сероводород, тиосульфат, дитионит, тетрасульфат, сульфит. Кроме того, Т. ferroxidans активно участвует в окислении сульфидов тяжелых металлов, которые нерастворимы в воде. К ним относятся такие минералы, как пирит (FeS2), халькопирит (CuFeS2), антимонит (SbS2), халькозин (Cu2S), кавеллин (CuS), пирротин (FeS), реальгар (AsS), виоларит (Ni2FeS4) и др. Остальные тионовые бактерии или такой возможностью не обладают, или она выражена слабо.
Отличительным свойством Т. ferroxidans является также способность окислять закисное железо в окисное. На этом основании данные бактерии могут рассматриваться как железобактерии, хотя относятся к роду Thiobacillus.
Как уже отмечалось выше, конечным продуктом окисления тионовыми бактериями молекулярной серы и различных ее соединений является сульфат. Если процесс идет таким образом, т.е. происходит полное окисление исходного субстрата, то результаты его отражают следующие уравнения. При окислении сероводорода:
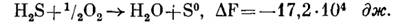
При окислении серы и тиосульфата в аэробных условиях:
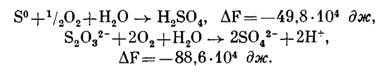
При окислении серы и тиосульфата Т. denitrificans в анаэробных условиях за счет использования нитратов:
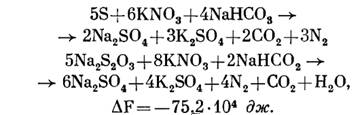
При окислении Т. thiocyanoxidans роданида:
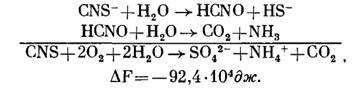
Однако нередко окисление идет не до конца, и в среде обнаруживаются различные не полностью окисленные продукты. Так при окислении сероводорода иногда появляется молекулярная сера, обнаружены также тиосульфат и политионаты. При окислении бактериями молекулярной серы констатировали появление тиосульфата и политионатов. Окисление тиосульфата также часто сопровождается образованием политионатов (тритионата, тетратионата, пеитатиопата) и элементарной серы. К числу не полностью окисленных продуктов относится и сульфит. Но не все из этих соединений могут являться результатом ферментативных процессов и действительно относятся к промежуточным продуктам окисления бактериями исходного субстрата. Некоторые из них образуются, видимо, чисто химическим путем или в результате побочных биологических реакций. Поэтому выяснение путей окисления микроорганизмами различных соединений серы весьма сложно, и далеко не все реакции до сих пор выяснены.
Очень мало еще данных о природе первого этапа превращения сульфида и не выяснено до конца, является сера прямым или побочным продуктом его окисления.
Весьма сложен вопрос об использовании бактериями элементарной серы, причем он имеет два аспекта: каким образом микроорганизмы воздействуют на это нерастворимое в воде вещество и как происходит сам процесс окисления.
В отношении механизма воздействия тиобацилл на серу существуют две точки зрения.
1. Для окисления бактериями серы необходим непосредственный контакт ее с клетками.
2. Сера используется бактериями после предварительного растворения в веществах липидной природы, выделяемых ими в среду.
При росте Т. thiooxidans на среде с молекулярной серой в среде обнаруживаются фосфолипиды. По одним данным тАУ это фосфатидилинозит, по другим тАУ несколько иные соединения (фосфатидил-N-метилэтанол, фосфатидилглицерин, дифосфатидилглицерин), причем их накопление совпадает с фазой активного роста культур. Тем не менее, считать доказанным второе предположение нельзя. Вполне вероятно, что для окисления серы бактериями важен и контакт ее с клетками, и выделение ими определенных веществ, которые ВлсмачиваютВ» серу.
Что касается пути окисления серы, то данные также довольно разноречивы. Наиболее вероятной считается следующая схема, которая приемлема также для окисления сероводорода:
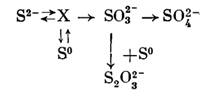
Предполагается, что X либо производное глутатиона, с которым реагирует сероводород или сера, либо тиол, связанный с мембранами клеток. Образование тиосульфата рассматривается как неферментативный процесс, который может идти без участия бактерий.
Большое число исследований посвящено изучению окисления тиосульфата. Данные опытов, проведенных с целыми клетками и бесклеточными препаратами разных видов тиобацилл, позволяют считать, что начальный зтап превращения тиосульфата может быть связан либо с его восстановлением под действием редуктазы с образованием сероводорода и сульфита:
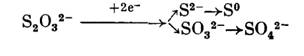
Либо с расщеплением до элементарной серы и сульфита:
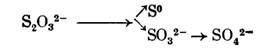
либо, наконец, с окислением до тетратионата и затем превращением в тритионат и сульфит:
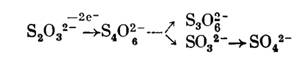
Предполагают, однако, что последний путь превращения тиосульфата не является основным. Что касается расщепления тиосульфата с образованием сульфита, то такую реакцию может осуществлять родоназа, связывающая серу в виде тиоцианата:
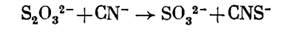
Она широко распространена не только у тионовых и других бактерий, но также у животных. Однако участвует ли этот фермент в метаболизме тиосульфата тионовыми бактериями, окончательно не установлено.
Значительно более исследован путь окисления сульфита. Работами, проведенными в лаборатории Пека с Т. thioparus, показано, что сульфит взаимодействует с аденозинмонофосфатом (АМФ), что приводит к образованию соединения, называемого аденилил сульфатом или аденозинфосфосульфатом (АФС). На следующем этапе происходит реакция между АФС и неорганическим фосфатом (Фн) с образованием аденозиндифосфата (АДФ) и освобождением свободного сульфата:

В результате действия фермента аденилаткиназы две молекулы АДФ могут превращаться в АТФ и снова давать АМФ:
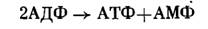
Таким образом, данный путь окисления сульфита связан с получением энергии (АТФ) в результате так называемого субстратного фосфорилирования. В то же время процесс окисления сульфита может сопровождаться передачей электронов в дыхательную цепь, функционирование которой сопряжено с синтезом АТФ.
Для ряда тионовых бактерий, в том числе Т. thioparus, показано, что окисление сульфита возможно и без образования АФС, в результате действия сульфитокисляющего фермента, который обеспечивает передачу электронов в дыхательную цепь на уровне цитохромов:
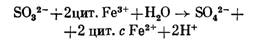
Не исключено, что у одного и того же организма могут функционировать разные пути окисления сульфита и других соединений серы, и значение того или иного зависит от условий среды и других факторов. На основании имеющихся к настоящему времени данных окисление тионовыми бактериями разных соединений серы можно представить следующей обобщенной схемой.
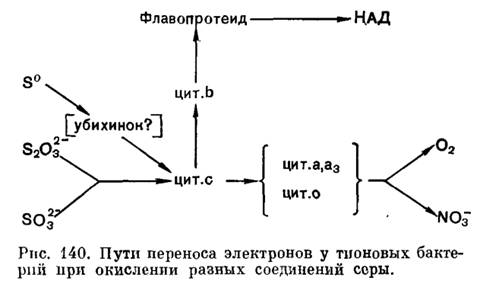
Что касается компонентов электрон-транспортных систем тионовых бактерий, то, по всем данным, в состав их у разных видов всегда входят цитохромы типа с. Обнаружены также цитохромы типа b и, видимо, его вариант, называемый цитохромом о, а у некоторых представителей тАУ цитохромы а и d. Кроме того, в электрон-транспортную систему входят, видимо, флавопротеиды и убихиноны. Но дыхательная цепь тионовых бактерий, обеспечивающая передачу электронов на кислород, как и у нитрифицирующих бактерий, сравнительно короткая, так как окисляемые субстраты имеют довольно высокий окислительно-восстановительный потенциал. Поэтому для данных микроорганизмов важное значение имеет энергозависимый перенос электрона против термодинамического градиента (обратный перепое электрона). Он обеспечивает образование восстановленного НАД, необходимого для ассимиляции углекислоты и других конструктивных процессов. Следует также отметить, что компоненты электронтранспортных систем и пути перепоса электронов не только могут иметь определеппые различия у разных представителей тиоповых бактерий, но и зависеть от характера окисляемого субстрата. Поэтому обобщенная схема (рис. 140) весьма условна.
По расчетам разных авторов, эффективность использования тионовыми бактериями свободной энергии составляет от 2 до 37%. Как и другие хемоавтотрофы, тионовые бактерии ассимилируют углекислоту в основном через цикл Кальвина. Но они обладают также способностью осуществлять другие реакции карбоксилирования, которые имеют важное значение для образования некоторых метаболитов.
Экстремально термоацидофильные серозависимые археи также способны к литотрофному росту в присутствии элементной серы и могут развиваться при 40тАУ100 В°C и рН 1тАУ6, это обитатели гидротерм, кальдер вулканов.
Гетеротрофные сероокисляющие микроорганизмы (организмы, использующие для питания органические вещества).
О способности некоторых облигатно-гетеротрофных бактерий, дрожжей и микромицетов окислять различные неорганические соединения серы известно довольно давно, хотя значение этих реакций в метаболизме гетеротрофов до сих пор до конца не выяснено. Среди гетеротрофных сероокисляющих бактерий преобладают виды группы псевдомонад, осуществляющие неполное окисление тиосульфата до тетратионата.
Еще С.Н. Виноградский отметил (1887тАУ1889), что бесцветные серобактерии могут расти в воде, содержащей очень небольшие количества органических веществ, и предполагал поэтому, что они способны усваивать углекислоту. На основании изучения физиологии разных штаммов Beggiatoa Прингсхейм считает, что среди них есть автотрофы, окисляющие сероводород и фиксирующие СО2, и есть представители, нуждающиеся в органических соединениях. Но и ряд гетеротрофных штаммов Beggiatoa в присутствии органических соединений окисляет сероводород, возможно, с получением энергии, т.е. они являются хемолитогетеротрофами. Наряду с бесцветными серобактериями известны типичные гетеротрофные микроорганизмы, участвующие в окислении сероводорода, молекулярной серы и тиосульфата. К числу таковых относятся представители Bacillus, Pseudomonas, Achromobacter, Sphaerotilus, а также актиномицетов, плесневых грибов (Penicillium luteum, Aspergillus niger), дрожжей и Alternaria. Некоторые из них, в частности нитчатая многоклеточная бактерия Sphaerotilus natans, в присутствии сероводорода откладывает в клетках серу. Другие (Pseudomonas aeruginosa, Ps. fluorescens, Achromobacter stuzeri) способны окислять тиосульфат до тетратионата (Na2S4O6). Отмечено также образование политионатов и сульфата при воздействии смешанных культур гетеротрофных микроорганизмов на элементарную серу. Механизм окисления и биологическое значение этого процесса для гетеротрофов остаются не выясненными. Некоторые гетеротрофные бесцветные серобактерии способны окислять соединения серы перекисью водорода, супероксидным радикалом, образующимся в клетках при окислении органических веществ: H2O2 + H2S → S0 + 2H2O. Физиологический смысл этого процесса тАУ детоксикация токсичных продуктов неполного восстановления кислорода (H2O2, O2-).
Полное окисление серных соединений до сульфата более характерно для микромицетов, обитающих в почвах, обогащенных элементной серой. Для некоторых видов родов микроскопических грибов Aspergillus, Penicillium, Trichoderma, Fusarium, Mucor и Auerobasidium показана способность к окислению элементной серы, тиосульфата и даже сульфидов металлов в сульфат, однако скорость такого окисления на один-два порядка ниже, чем у литотрофных бактерий.
сера циклический превращение микроорганизм
Заключение
В последние десятилетия природный цикл серы подвергается усиливающемуся антропогенному воздействию, приводя к накоплению токсических соединений серы и нарушению баланса природного цикла серы. В частности, в результате крупномасштабных выбросов серных соединений образуются двуокись серы, выделяемая ТЭЦ при сжигании органического топлива, сероводород и летучие органические сульфиды, выделяемые целлюлозно-бумажными и металлургическими предприятиями, а также при разложении муниципальных и сельскохозяйственных стоков. Эти соединения токсичны уже в микрограммовых концентрациях. Они способны отравлять воздух, влиять на атмосферную химию, вызывать дефицит растворенного в воде кислорода.
Поэтому использование прокариот, участвующих в превращении соединений серы, детоксикации токсичных соединений серы, представляется весьма актуальным. Кроме того, чрезвычайно высокая каталитическая активность микроорганизмов является причиной того, что они играют главную роль в химических превращениях, происходящих на поверхности Земли. Благодаря небольшим размерам микроорганизмы обладают по сравнению с животными и высшими растениями высоким соотношением поверхности и объема, что и приводит к быстрому обмену субстратов и продуктов выделения между клеткой и окружающей средой.
Важными факторами являются также высокая скорость их размножения в благоприятных условиях и широкая распространенность по всей биосфере. Однако, несмотря на исключительное значение прокариот в трансформации биогенных элементов, до сих пор масштабы деятельности прокариот в круговороте веществ, и в частности в цикле серы, до конца не оценены.
Литература
1. Грабович М.Ю. Участие прокариот в круговороте серы // Соросовский Образовательный Журнал, 1999, №12, с. 16тАУ20.
2. Громов Б.В., Павленко Г.В. Экология бактерий. Л.: Изд-во ЛГУ, 1989. 248 c.
3. Громов Б.В. Удивительный мир архей // Соросовский Образовательный Журнал. 1997. №4. C. 23тАУ26.
4. Кальдерные микроорганизмы / Под ред. Г.А. Заварзина. M.: Наука, 1989. 120 c.
5. Кондратьева Е.Н. Хемолитотрофы и метилотрофы. М.: Изд-во МГУ, 1983. 172 c.
6. Малахов В.В. Вестиментиферы тАУ автотрофные животные // Соросовский Образовательный Журнал. 1997. №9. C. 18тАУ26.
Вместе с этим смотрят:
Анатомия и физиология заднего мозга. Строение и механизм кровообращения
Бiологiчне рiзноманiття людських рас
Бiологiя iндивiдуального розвитку
Бiологiя лева
Банан